БИОХИМИЯ
УДК 577.612.01
Р. М. Симонян, Г. М. Симонян, М. А. Симонян
Ингибирование цитохромом b558III супероксид-продуцирующей
активности супрола и ее стимулирование
тетрахлором углерода in vitro
(Представлено академиком М. А. Давтянoм 16/III 2005)
Супероксид-продуцирующий липопротеин - супрол
локализован в сыворотке крови млекопитающих [1] и рыб [2] и является
липопротеином высокой плотности, продуцирующим супероксидные радикалы
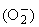 в присутствии следов ионов
переходных металлов (Fe+3, Cu+2) in vitro и in vivo [3].
Это имеет место, в частности, при интоксикации крыс ионами переходных металлов
[4,5], а также при сердечно-сосудистых заболеваниях [6]. Продуцированные
в присутствии следов ионов
переходных металлов (Fe+3, Cu+2) in vitro и in vivo [3].
Это имеет место, в частности, при интоксикации крыс ионами переходных металлов
[4,5], а также при сердечно-сосудистых заболеваниях [6]. Продуцированные
 и их производные
(1O2, H2O2, HO·, NO· и др.)
инициируют перекисное окисление фосфолипидных остатков самого супрола, вызывая
агрегацию супрола. Это, в свою очередь, может изменить вязкость крови и
воздействовать на гемодинамику и тромбообразование [7]. Супрол продуцирует
в течение сравнительно
долгого времени (5-6 ч) в аэробных условиях in vitro, в отличие от других
-продуцирующих систем [7]. При
этом продуцированные
нормально дисмутируются антиоксидантными препаратами различного характера [7].
Обнаружение биологических средств, ингибирующих продуцирование
супролом, может иметь важное
значение для стабилизации или предотвращения агрерации супрола. По
предварительным результатам супрол продуцирует
и в присутствии следов "связанного" железа (например,
железа гемовой группы гемоглобина), а цитохром (цит) b558III обладает
способностью образовывать нестабильный комплекс с метгемоглобином (метHb) за
счет NO·, присутствующего в гемовой группе
цит b558III как 5-й или 6-й лиганд железа [8]. Целью работы явилось определение
характера воздействия цит b558III и CCl4 на супероксид-продуцирующую
активность супрола in vitro
и их производные
(1O2, H2O2, HO·, NO· и др.)
инициируют перекисное окисление фосфолипидных остатков самого супрола, вызывая
агрегацию супрола. Это, в свою очередь, может изменить вязкость крови и
воздействовать на гемодинамику и тромбообразование [7]. Супрол продуцирует
в течение сравнительно
долгого времени (5-6 ч) в аэробных условиях in vitro, в отличие от других
-продуцирующих систем [7]. При
этом продуцированные
нормально дисмутируются антиоксидантными препаратами различного характера [7].
Обнаружение биологических средств, ингибирующих продуцирование
супролом, может иметь важное
значение для стабилизации или предотвращения агрерации супрола. По
предварительным результатам супрол продуцирует
и в присутствии следов "связанного" железа (например,
железа гемовой группы гемоглобина), а цитохром (цит) b558III обладает
способностью образовывать нестабильный комплекс с метгемоглобином (метHb) за
счет NO·, присутствующего в гемовой группе
цит b558III как 5-й или 6-й лиганд железа [8]. Целью работы явилось определение
характера воздействия цит b558III и CCl4 на супероксид-продуцирующую
активность супрола in vitro
Цит b558III из
эритроцитарных мембран (ЭМ) и супрол из сыворотки крови крыс получали
биотехнологическим способом [9], без использования детергента [10]. При этом цит
b558III получается в электрофоретически гомогенном состоянии, а супрол содержит
небольшие следы гемопротеина (Hb). Активирование супрола (для продуцирования
) осуществляли следами ионов
Fe+3 [3].
-продуцирующую активность супрола и цит b558III
определяли нитротетразолиевым синим (НТС) методом [9], рассчитав проценты
повышения плотности максимального оптического поглощения формазана (при 560 нм),
образовавшегося в результате восстановления НТС супероксидными радикалами. За
единицу
- продуцирующей
активности супрола или цит b558III принимали количество белка, вызывающее
50%-ное увеличение плотности поглощения формазана. Удельная
- продуцирующая активность
супрола и цит b558III была определена в расчете на 1 мл сыворотки и эритроцитов
соответственно.
Оптические спектры
поглощения регистрировали на спектрофотометре ''Specord UV/VIS'' (Германия) с
длиной оптического пути 1 см при 20o.
Статистическую обработку полученных результатов осуществляли методом
вариационной статистики Стьюдента-Фишера, с расчетом критерия достоверности
P.
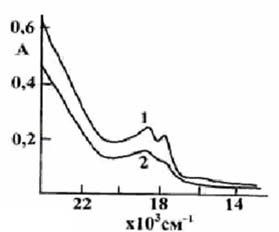
В результате небольшого гемолиза
эритроцитов (наблюдающегося при декапитировании крыс) супрол образует
нестабильное комплексное соединение с метHb (рис.1,1). После активирования
такого супрола ионами Fe+3 форма его оптического спектра не
изменяется. При этом на спектре видны характерные максимальные поглощения Hb
(альфа- и бетта-полосы поглощения).
|
|
Рис.1.
После агрегации (самоосаждения)
супрола в результате инкубирования при 4o в течение 40-50 ч в
аэробных условиях и центрифугирования его осадoк имеет красноватый цвет, а
супернатант бесцветный. Это свидетельствует о том, что Hb действительно входит в
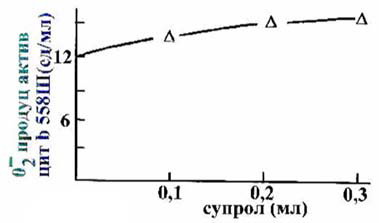
комплексное соединение с супролом. После гомогенизации в 0.04 М калий фосфатном
буфере (КФБ) осадка агрегированного с Hb супрола смесь проявляет
- продуцирующую активность
(рис.2). Под влиянием добавленного цит b558III (A530 = 0.2) наблюдается
снижение (60.5±6.4%, n = 6, P < 0.05)
- продуцирующей активности
супрола с A430 = 0.02 в реакционной смеси. В результате его
инкубирования (в течение 20-30 мин) с цит b558III при 20o наблюдается
отщепление Hb от супрола и присоединение его к цит b558III.
Рис.2.
При этом осадок агрегированного
супрола теряет красный цвет, а супернатант (комплекс цит b558III с Hb)
представляет из себя раствор красноватого цвета. Спектр этого раствора имеет
смешанную форму спектров Hb и цит b558III, что харктерно для комплекса Hb с цит
b558III [11] (рис.3).
Рис.3.
НАДРН-зависимая
-продуцирующая активность
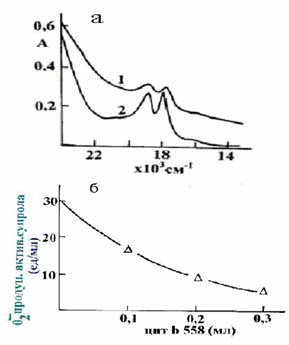
комплекса цит b558III с Нb на 20-30% выше таковой у цит b558III (рис.4, а).
Bидимо, в этом состоянии Hb, как ионы переходных металлов, служит переносчиком
электрона от НАДРН (группа в молекуле супрола) к молекулярному кислороду,
превращая его в
[3]. Цит
b558III входит в комплексное соединение с отщепленным от супрола Hb, в
результате чего происходит дезактивирование супрола. Одновременно несколько
повышается
-продуцирующая
активность цит b558III. Фактически цит b558III очищает супрол от следов Hb и
дезактивирyет этот липопротеин сыворотки высокой плотности, повышая его
стабильность. Таким образом, проявляется "антиоксидантный" эффект цит b558III,
предотвращающий продуцирование
супролом и подавляющий перекисное окисление
фосфолипидных остатков супрола супероксидными радикалами. Это имеет большое
значение для сохранения стабильности супрола, соответственно и нормальной
вязкости сыворотки.
Рис.4.
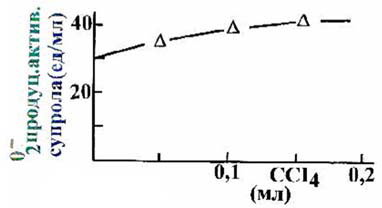
Под
воздействием CCl4 (рис 4, б) наблюдается повышение
- продуцирующей активности
супрола (31.4±3.2%, P < 0.03), что, возможно,
связано с повышением "растворимости" супрола как липопротеина высокой плотности
в гидрофобной среде CCl4. Видимо, на начальных этапах
экспериментального цирроза печени крыс под воздействием CCl4
происходит повышение
-продуцирующей активности и липидной пероксидации
супрола с соответственным повышением вязкости сыворотки и возможным изменением
гемодинамики. Эти обстоятельства вынуждают переоценить токсические эффекты
CCl4. С другой стороны, подавление
-продуцирующей активности супрола, расщепление
комплекса супрола с Нb и захват последнего цитохромом b558III являются новыми
факторами "очищения" сыворотки, повышения стабильности липопротеина сыворотки
высокой плотности - супрола и сохранения нормальной вязкости крови в целом.
Исходя из того, что цит b558III локализован на внешней поверхности ЭМ [11], не
исключается аналогичная роль цит b558III и в гетерогенной фазе (в ЭМ).
Институт биохимии им. Г. X. Бунятяна НАН РА
Литература
1. Симонян М. А., Карапетян А. В., Бабаян М.
А., Симонян М. А. - Биохимия. 1996. Т. 61. № 5. С. 932-938.
2. Симонян Г. М., Симонян М. А., Карагезян
К. Г. - ДНАН Армении. 2003. Т. 103. № 2. С. 151-153.
3. Симонян Г. М., Бабаян М. А., Симонян Р.
М., Симонян М. А. - Биол. ж. Армении. 1999. Т. 1. № 52. С.
18-21.
4. Оксузян Г. Р., Симонян Р. М.,
Симонян М. А., Алексанян С. С. - Мед. наука Армении. 2001. Т. 41.
№ 2. С. 21-26.
5. Оксузян Г. Р., Симонян Р.
М., Бабаян М. А., Симонян М. А. - Мед. наука Армении. 2002. T. 42.
№ 2. С. 3-6.
6. Алексанян Серг. С.,
Алексанян С. С., Симонян Г. М. - Вестник МАНЭБ Санкт-Петербурга.
2004. Т. 9. № 8. С. 198-200.
7. Симонян Р.
М., Симонян Г. М., Симонян М. А. - Вестник МАНЭБ Санкт-Петербурга.
2004. Т. 9. № 8. С. 72-79.
8. Симонян Р. М. Новые факторы оксидативного повреждения компонентов крови.
Автореф. канд. дис. Ин-т биохимии НАН РА. Ереван. 2004.
9. Симонян М. А., Симонян Г. М. Способ получения
металлопротеинов крови. Лицензия изобрет. № 341 Армпатента. Ереван. 1997.
10. Симонян М. А., Симонян Г. М., Симонян
Р. М. Способ получения цитохромов b из мембран эритроцитов.
Лицензия № 908 Армпатента. Ереван. 2001.
11.
Симонян Г. М., Симонян Р. М., Бабаян М. А., Карапетян А. В., Симонян М. А. -
Мед. наука Армении. 2003. Т. 43, № 1. С. 30-38.