фосфолипидов в мозговой ткани при моделированном коразолом
эпилептиформном припадке
|
|
УДК 616.36.004
Oсобенности качественно-количественных изменений
фосфолипидов
в мозговой ткани белых крыс при эпилептиформных припадках и на
фоне
предварительной сенсибилизации их факторами
антиоксидантного действия
(Представлено 22/VII 2004)
Моделирование эпилептиформных припадков,
индуцированных коразолом, сопровождается чувствительными нарушениями
стабилизированного в норме статуса фосфолипид-фосфолипидных1 соотношений в мозговом веществе
экспериментальных белых крыс [1]. Постоянству качественного набора и
количественного содержания филогенетически запрограммированного стереотипа ФЛ в
различных биологических системах организма отводится принципиально важная роль в
обеспечении норм клеточной активности [2,3]. Исходя из этого становится
очевидной необратимость выраженных расстройств в физиологически существующем
динамическом равновесии между системами про- и антиоксидантного действия,
завершающихся гибелью организма. Таблица 1
Мы задались
целью проследить за характером сдвигов качественно-количественного состава всего
спектра ФЛ в мозговой ткани экспериментальных белых крыс при коразоловых
припадках, вызванных как у интактных животных, так и спустя 10-15 мин после
предварительного введения им тиосульфата натрия и витамина Е. Такая постановка
вопроса имеет принципиально важное значение, поскольку ТСН в настоящее время
отводится роль мощного синергиста гидроксической формы витЕ как единственной в
проявлении антиоксидантных свойств в сложной эндогенной системе антирадикальной
защиты клетки. Этот факт был зарегистрирован при многочисленных болезненных
состояниях организма - отравлениях различного происхождения [4-8], токсикозах
анестетического действия [9], сахарном диабете
[10-14].
Исследования проведены на 50
беспородных белых крысах-самцах массой 180-200 г, содержавшихся в обычных
условиях вивария. Одноразовое введение коразола производилось внутримышечно из
расчета 8-9 мг/весь вес, ТСН - 1 мг/весь вес и витЕ - 0.4 мг/весь вес. При
формировании коразоловых припадков на фоне предварительного введения ТСН и витЕ
инъекции коразола производили спустя 15 мин после введения указанных препаратов
антиоксидантного действия. Животных умерщвляли декапитированием;
стабилизированную оксалатом цельную кровь (в соотношении 1:9), мозговую ткань,
освобожденную от оболочек и кровеносных сосудов, хранили в условиях
холода.
Экстракцию ФЛ производили методом
Фолча [15] из предварительно полученных ацетоновых порошков исследуемого
материала в модификации Карагезяна [16]. Количество ФЛ рассчитывали в мкг
минерализованного липидного фосфора на г сухого остатка исследуемого материала
или на мг белка, определенного по Лоури [17], и выражали в процентах от суммы
всех
ФЛ.
|
|
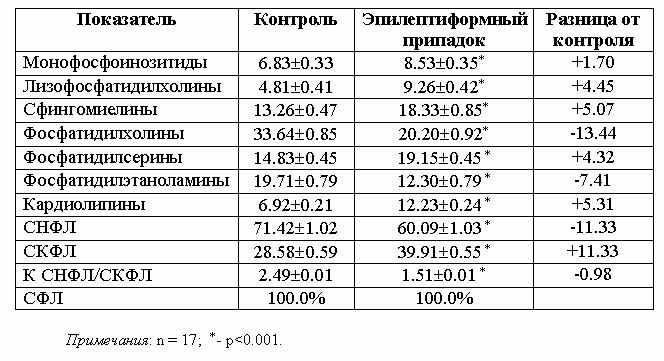
Согласно результатам первой серии
исследований (табл.1) развитие коразоловых припадков характеризуется ярко
проявляющимися расстройствами в картине качественно-количественных сдвигов
нейтральных и кислых представителей ФЛ в мозговой ткани экспериментальных
животных. Очевидно статистически достоверное увеличение в мозговом веществе под
действием одного только коразола количественного содержания всех индивидуальных
представителей КФЛ - МФИ, ФС и КЛ. Эти сдвиги сопровождаются параллельно
развивающимися разнонаправленными отклонениями уровня отдельных фракций НФЛ.
Примечательно, что наблюдающееся при этом резко выраженное уменьшение
фосфатидилхолинов сопровождается возрастанием количества лизофосфатидилхолинов,
что является результатом интенсивно совершающегося деацилирования ФХ,
катализируемого чрезмерно активировавшейся фосфолипазой A2.
Наблюдающееся при этом уменьшение уровня фосфатидилэтаноламинов интерпретируется
как результат активирующихся при патологических состояниях, особенно при
поражениях ЦНС, процессов деметилирования, сопровождающихся в данном случае
чрезмерным возрастанием количества ФХ, интенсивно деацилирующихся, как
отмечалось выше, под действием ФЛазы A2. Таким образом, содержание ФХ
не только не возрастает, а наоборот, даже заметно убывает по сравнению с
исходными величинами. Особого внимания заслуживает чувствительное увеличение в
мозговой ткани при коразоловом поражении содержания всех представителей КФЛ -
МФИ, ФС и КЛ, имеющих прямое отношение к процессам тканевого дыхания [18],
заметно подавляющегося при изученной патологии, в качестве
компенсаторно-приспособительных факторов.
Анализ полученных данных свидетельствует о том, что в результате
описанных превращений в метаболизме мозговых ФЛ имеет место заметное уменьшение
СНФЛ при одновременно наблюдающемся возрастании процентного содержания СКФЛ в
общей СФЛ, что отражается на величине К СНФЛ/СКФЛ, резко уменьшающейся при
данной патологии.
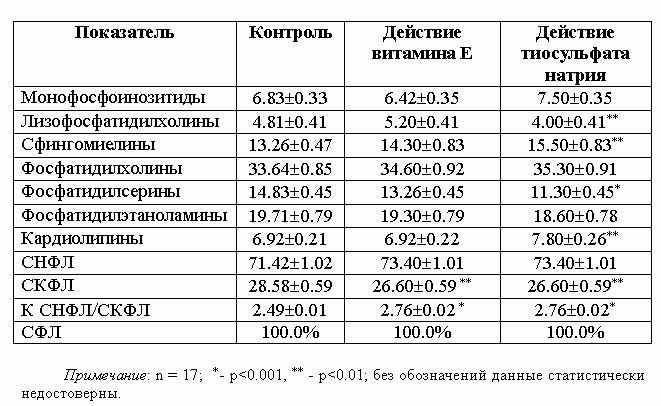
В следующей серии
экспериментов исследовалось воздействие изолированно примененных ТСН и витЕ на
метаболизм ФЛ мозговой ткани интактных белых крыс. Как вытекает из данных табл.
2, ТСН и витЕ в отдельности не вызывают существенных отклонений в метаболизме ФЛ
мозговой ткани интактных животных, несмотря на то, что даже незначительные
отклонения в СНФЛ и СКФЛ влияют на величину К
СНФЛ/СКФЛ.
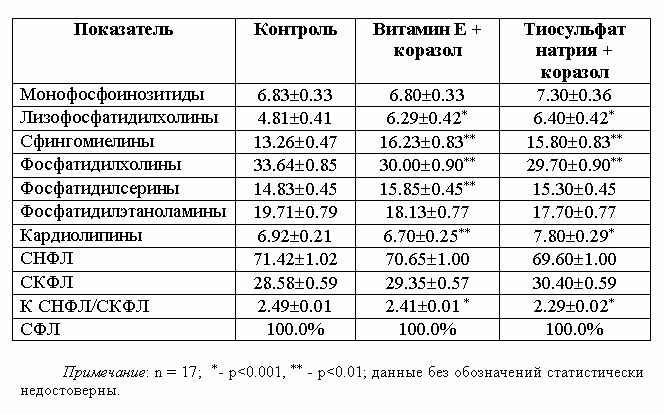
В табл. 3 приведены данные об
изменениях метаболизма ФЛ в мозговой ткани при коразоловых приступах, вызванных
на фоне предварительно введенных витЕ и ТСН.
Таблица 2
|
|
Таблица 3
|
|
Коразоловый припадок на фоне предварительно
введенных за 15 мин растворов ТСН и витЕ характеризуется слабо выраженными
отклонениями всех изученных показателей метаболизма ФЛ в мозговой ткани
экспериментальных животных. Исключением в обоих вариантах эксперимента является
сохраняющееся доминирование количественного содержания ЛФХ над контрольными
показателями, сопровождающееся одновременно убылью содержания ФХ, что указывает
на высокую степень активности ФЛазы А2.
Таким образом, результаты проведенных исследований свидетельствуют о
важной нивелирующей роли препаратов антиоксидантного действия - витЕ и ТСН в
поддержании нормального физиологического статуса функционирования реакций
метаболизма ФЛ в мозговой ткани белых крыс с моделированным коразолом
эпилептиформным припадком.
Институт молекулярной биологии НАН
РА
Институт биохимии им. Г. Х. Бунятяна НАН
РА
1. Мартиросян М. А. Некоторые показатели липидного обмена при коразоловых приступах. Канд.
дис. Ереван. 1990. 108 с. 1ВитЕ - витамин Е, К - коэффициент, КЛ - кардиолипины, КФЛ - кислые
фосфолипиды, ЛФХ - лизофосфатидилхолины, МФИ - монофосфоинозитиды, НФЛ -
нейтральные фосфолипиды, СКФЛ - сумма КФЛ, СНФЛ - сумма НФЛ, СФЛ - сумма всех
ФЛ, ТСН - тиосульфат натрия, ФЛ - фосфолипиды, ФЛазаА2 - фосфолипаза
A2, ФС - фосфатидилсерин, ФХ - фосфатидилхолин, ФЭ -
фосфатидилэтаноламин.
2. Крепс Е. М. - XXII Баховские чтения. Л. Наука. 1967. С.
74.
3. Крепс Е. М. Липиды клеточных мембран. Л. Наука. 1981. 330 с.
4. Карагезян М. К., Бояджян А.
С., Осипян Л. Л., Карагезян К. Г. - Тезисы докл. V съезда Арм.
физиол. о-ва. Ереван. 1994. С. 44.
5. Карагезян М. К., Бояджян А. С., Осипян Л. Л., Карагезян К. Г. - Докл. РАН. 1995. Т. 341. №1. С. 113-114.
6. Карагезян М. К., Овсепян Л. М., Овакимян С. С., Бояджян
А. С., Осипян Л. Л., Карагезян К. Г. - Докл. РАН. 1995. Т. 341.
№2. С. 259-262.
7. Карагезян
М. К., Овакимян С. С., Овсепян Л. М., Бояджян А. С., Осипян Л. Л., Карагезян К.
Г. - Докл. РАН. 1995. Т. 341. №3. С.
408-411.
8. Карагезян М. К. Изучение молекулярных механизмов токсических эффектов микотоксина
зеараленона. Канд. дис. Ереван. 1997. 120 с.
9. Данилова Р. Л. Действие галотановой
анестезии на мембранные структуры мозговой ткани и эритроцитов и методы его
коррекции. Автореф. канд. дис. 2000. 20 с.
10. Едоян А. Р. Специфика корригирующего
действия сверхнизких доз факторов химической и физической природы при нарушениях
метаболизма фосфолипидов у белых крыс с моделированным аллоксаном сахарным
диабетом. Автореф. канд. дис. Ереван. 2004. 21 с.
11. Едоян Л. В., Карян Ш. С., Едоян А. Р., Карагезян К. Г.
- Информационные технологии и управление. Ереван. 2003. Т. 2. С.
212-218.
12. Карян Ш. С. Особенности антирадикального действия сверхнизких доз физиологически
активных соединений и лазерного облучения при аллоксановом диабете. Автореф.
канд. дис. Ереван. 2004. 23 с.
13. Мартиросян Э. А. Изучение гликосфинголипидов и сфингозина
в печени и сердце белых крыс при аллоксановом диабете и при его лечении.
Автореф. канд. дис. Ереван. 2004. 20 с.
14.
Геворкян Д. М. Липидный обмен при сахарном диабете и
коррекция его антиоксидантами. Автореф. канд. дис. Ереван. 1989. 45
с.
15. Folch J., Lees M.,
Sloane-Stane G. - J. Biol. Chem. 1957. V. 226. P. 497-509.
16. Карагезян К. Г. Роль фосфолипидов в жизнедеятельности организма. Ереван. 1972. 267 с.
17. Lowry O. H., Rosebrough N.
J., Farr A. L et al. - J. Biol Chem. 1951. V. 193. №1. P. 265-275.
18. Бурлакова Е. Б., Джалябова
М. И., Гвахария В. О., Глущенко Н. Н., Молочкина Е. М., Штолько В. Н. В кн.: Биоантиокислители в регуляции метаболизма в норме и патологии. М.
Наука. 1982. С. 113-140.
Footnotes: