ФИЗИОЛОГИЯ
УДК 612.826+612.886
Л. П. Манукян, В. А. Саргсян
Синаптические механизмы вестибуло-таламических
взаимоотношений
(Представлено чл.-кор. НАН РА Л. Р. Манвеляном
9/III 2004)
Известно, что таламус является основным реле
передачи восходящих вестибулярных сигналов в кору мозга [1-3]. Основным
носителем этих сигналов является восходящий тракт Дейтерса , который хорошо
представлен почти у всех млекопитающих [4]. Проекции в составе этого тракта
достигают коры мозга обезьяны через вентрозаднее ядро таламуса [5,6], а также
зрительной коры кошки через центральное латеральное и парацентральное ядра
таламуса [7]. С целью завершения серии наблюдений по афферентному контролю
активности вестибуло-спинальных (ВС) нейронов ядра Дейтерса [8] была проведена
настоящая работа по исследованию антидромных и синаптических механизмов
вестибуло-таламических взаимоотношений.
Опыты проведены на 57 взрослых кошках массой
2.5-3.5 кг, наркотизированных нембуталом (45-50 мг/кг) или 2% рометаром или
калипсолом (0.5 мг/кг). Биполярными вольфрамовыми электродами раздражались
таламическое ядро Centrum Medianum (билатерально) (СМ) и вентрозаднее ядро
таламуса (VPL) (контралатерально), соответственно по следующим стереотаксическим
координатам: L 3, H 1, A 7 и L 6, H 1, A 9 [9]. ВС нейроны ядра Дейтерса
идентифицировались по их антидромным потенциалaм действия (ПД), вызванным
раздражением латерального ВС тракта на уровне С2-3 шейных сегментов спинного мозга. Биполярно
раздражались вестибулярные нервы обеих сторон (0.1-0.15 мА, 0.1-0.5 мс, 1.0-10.0
В). Внутриклеточная активность нейронов ядра Дейтерса регистрировалась
стеклянными микроэлектродами с сопротивлением 2-10 МOм, заполненными 2М
раствором KCl. Точки раздражения и отведения верифицировались гистологически.
Зарегистрирована внутриклеточная активность
292 нейронов латерального вестибуярного ядра (ЛВЯ), из них 154 реагировали на
раздражение СМ и VPL
таламуса.
|
|
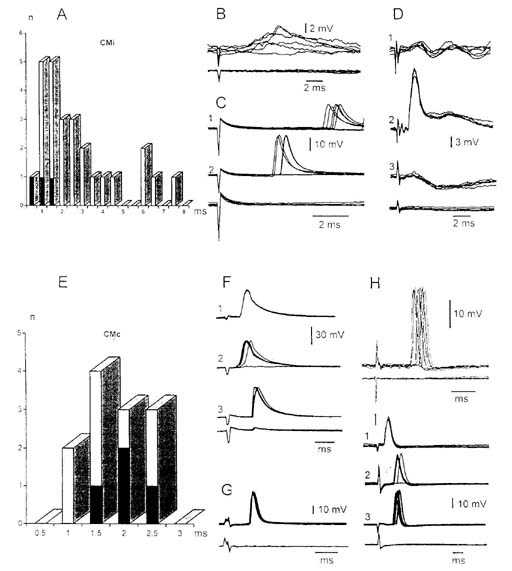
Рис. 1. Антидромные и синаптические потенциалы нейронов
ядра Дейтерса, вызванных
раздражением ипси- (СМi) и контралатерального (СМc)
ядер Centrum medianum таламуса.
А, E - гистограмма распределения скрытых
периодов потенциалов нейронов ядра Дейтерса
вызванные раздражением
соответственно СМi и СМc. Темные столбики на А, Е -
идентифицированные ВС
нейроны; В, D1 - ВПСП и ТПСП двух нейронов ядра Дейтерса,
вызванные
раздражением СМi; D2,3; F2 - конвергенция влияний из
ипси-(D 2, F 2) и
контралатерального (D 3) VIII нервов на нейроны ядра Дейтерса;
С1,2 - полисинаптические
ПД нейрона ядра Дейтерса при пороговом
(С1) и надпороговом (С2) раздражении; F 3, G -
антидромные
ПД двух нейронов, вызванные раздражением СМc; H - синаптический ПД
нейрона ядра
Дейтерса, вызванный раздражением СМc. F1, I1 - антидромное
ПД двух ВС
нейронов ядра Дейтерса, вызванное раздражением латерального ВС
тракта. F3 -
антидромный ПД того же нейрона на раздражение CMc.
I2,3 - конвергенция влияний из
CMc (I2) и VPL
(I3) на ВС нейрон ядра Дейтерса. Здесь и на следующем рисунке
приведенные потенциалы зарегистрированы усилителем постоянного тока при
наложении
5-10 пробегов луча; частота повторения 5/с. Нижние записи на В - I -
потенциалы поля при
внеклеточном отведении.
Ответы на раздражение
СМ. Раздражение как ипсилатерального СМ (СМi), так и контралатерального
СМ (СМc) вызвало в нейронах ядра Дейтерса преимущественно синаптические ответы
(рис.1,C,H). В случае раздражения СМi они возникали со скрытым периодом 0.8-26.0
(n=23) (рис.1,A). Моно- и полисинаптические возбуждающие постсинаптические
потенциалы (ВПСП) и тормозной постсинаптический потенциал (ТПСП) на раздражение
СМi регистрировались относительно реже, что, вероятно, можно объяснить низким
порогом деполяризации для вызова ортодромного потенциала действия (ПД) (рис.1,В
и D1). 5 нейронов из 23, активированных от СМi, были идентифицированы
как ВС нейроны (pис.1,А, темные столбики). В трех нейронах ядра Дейтерса
стимуляция СМi вызывала антидромные ПД.
Раздражение СМc вызывало в нейронах ядра
Дейтерса также преимущественно ортодромные ПД, со скрытым периодом 1.1-2.7
(рис.1,E). 4 из них были идентифицированы как ВС нейрoны (рис.1, E, темные
столбики; F3). Среди ортодромных ПД два возникали моносинаптически, а
остальные - ди- и полисинаптически. В четырех нейронах раздражение СМc вызывало
антидромные ПД (рис.1,F1,G). ВПСП и ТПСП в ответ на раздражение СМc
не наблюдались. Былa отмеченa конвергенция влияния от СМi, контра- и
ипсилатерального VIII нервов (рис.1,D), а также ипсилатерального VIII нерва и
СМc на ВС нейроны ядра Дейтерса (рис.1,F). В некоторых ВС нейронах ядра Дейтерса
наблюдалась конвергенция влияния от СМi и СМc (рис.1,I).
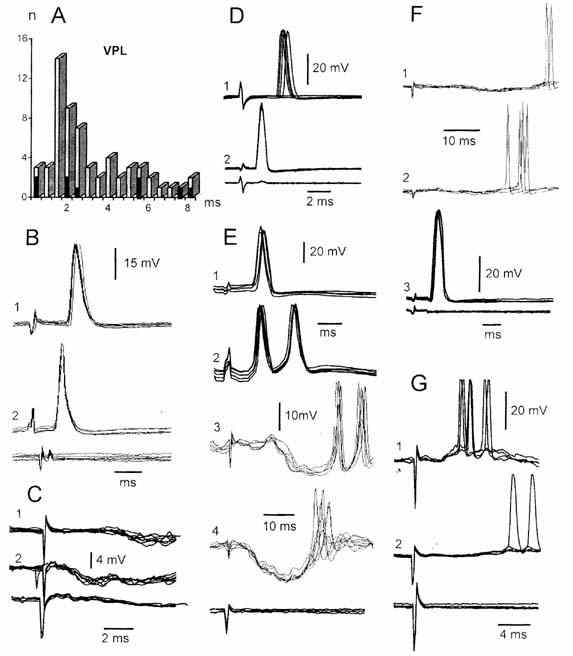
Ответы на раздражение VPL
таламуса. Раздражение контралатерального VPL таламуса в 108 нейронах ЛВЯ
вызывало в основном полисинаптические реакции. Они возникали со скрытым периодом
в широких пределах от 1.2 до 36.0 (pис.2,А). 22 нейрона из этой группы были
идентифицированы как ВС нейроны (pис.2,А темные столбики; В,2,
Е1,F3). В 8 нейронах вызывались полисинаптические ВПСП, на
верхушке которых, как правило, возникали вспышки ПД
(pис.2,F1,G1,2). Реже раздражение VPL таламуса вызывало
ТПСП
Рис. 2. - Антидромные и синаптические потенциалы нейронов
ядра Дейтерса, вызванные
раздражением контралатерального вентрозаднего ядра
таламуса. А - гистограмма
распределения скрытых периодов потенциалов нейронов
ядра Дейтерса вызванные
раздражением VPL. Темные столбики - идентифицированные
ВС нейроны. В1, D1, F1 и G -
ди-
(В1) и полисинаптические ПД трех нейронов ядра Дейтерса на
раздражение VPL; В2, D2,
Е1 и F3 -
антидромные ПД четырех ВС нейронов ядра Дейтерса, вызванные раздражением
латерального ВС тракта; С1, Е3 - ТПСП (С1) с
реакцией облегчения (Е3) двух нейронов ядра
Дейтерса на раздражение
VPL. С2, Е4 и F2 - ТПСП (С2,
Е4) и полисинаптические ПД двух
нейронов ядра Дейтерса на раздражение
контралатерального VIII нерва; Е2 - ответ нейрона
на раздражение
ипсилатерального VIII нерва; Е - пример конвергенции влияний из ипси-
(Е2) и контралатерального (Е4) VIII нервов и VPL
(Е3) на ВС нейрон (Е1) ядра Дейтерса.
Нижние записи на В -
G - потенциалы поля при внеклеточном отведении.
(pис.2,С1), которые завершались реакцией облегчения
(pис.2,Е3). В пяти нейронах ядра Дейтерса раздражение VPL таламуса
вызывало ПД с очень коротким скрытым периодом (0.1-0.3), которые, вероятно,
можно отнести к разряду интрааксональных. Наблюдалась широкая конвергенция
влияний на нейронах ядра Дейтерса со стороны VPL таламуса, контра- и
ипсилатерального VIII нервов (pис.2,Е).
Проведенные эксперименты показали, что
раздражение таламических ядер СМ и VPL в большинстве нейронов ядра Дейтерса
вызывает полисинаптические и реже моносинаптические реакции. Это свидетельствует
о том, что, как и в случае множества уже изученных структур ЦНС [8,10], таламус
также прямо или опосредованно модулирует деятельность ВС системы. Регистрация
антидромных реакций нейронов ядра Дейтерса на раздражение СМ указывает также на
наличие обратной связи с таламусом. Полученная в таком случае вестибулярная
информация достигает затем коры мозга и дальше через кортико-спинальный путь
участвует в контроле мотонейронов спинного мозга.
Таким образом, вестибуло-таламические пути,
вовлекаясь в интегративную деятельность ядра Дейтерса, активно участвуют в
процессе регуляции позы и ориентации тела в пространстве.
Авторы выражают свою благодарность NFSAT за
содействие в приобретении установки ÄPP-1 all Purpose Pipette Puller".
Изготовленные на этой установке микроэлектроды использовались в проведении
настоящего исследования.
Институт физиологии им. Л. А. Орбели НАН РА
Литература
1. Maciewicz R. J., Phipps B.
S., Bry J., Highstein M. - Brain Res. 1982. V. 252. P.
1-11.
2. Nagata S. - Brain Res. 1986. V. 376. P. 57-70.
3. Wild J. M. - J. Comp. Neurol. 1988. V.
271. P. 451-460.
4. Baker R.,
Highstein S. M. - J. Neurophysiol. 1978. V. 41. P. 1929-1946.
5. Buttner U., Lang W. - In: R. Granit and O. Pompeiano, Progress in Brain Research. V. 50.
Reflex Control of Posture and Movement. Elsevier. Amsterdam. 1979. P. 581-588.
6. Buttner U., Henn V., Oswald
H. P. - Brain Res. 1977. V. 30. P.
435-444.
7. Vanni-Mercier G.,
Magnin M. - Exp. Brain Res. 1982. V.45. P. 451-455.
8. Sarkisian V. H. - Arch. Italian Biology. 2000. V. 138. P.
245-353.
9. Berman A. L. The brain stem of the cat. A cytoarchitectonic atlas with stereotaxic
coordinates. Wisconsin. Madison etc. 1968.
10. Wilson V. J., Melvill-Jones, G. - Mammalian Vestibular Physiology. Plenum Press. New York. London. 1979.
P. 356.