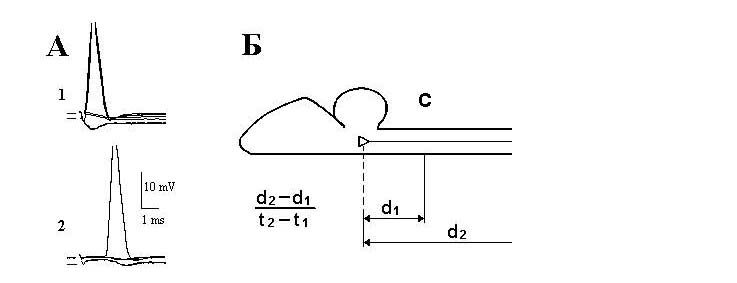
 |
лягушки. А - антидромные потенциалы действия С (1) и L (2) нейронов. Нижние кривые -
внеклеточные потенциалы поля после выхода микроэлектрода из клетки. Б - схема измерения
скорости проведения возбуждения по волокнам ретикулоспинального тракта; d1 и d2 - расстояния,
измеренные по вентральной поверхности спинного и продолговатого мозга от участков раздражения
шейных (С) и поясничных (L) сегментов до места входа микроэлектрода в мозг: t1 и t2 - скрытые
периоды антидромных ответов нейронов на стимуляцию шейных и поясничных сегментов спинного
мозга, соответственно.
периодов нейрона на раздражение шейного (С) и поясничного (L) сегментов спинного мозга (рис.1,Б), d1 и d2 соответствуют расстоянию между местом введения микроэлектрода в ствол мозга и участками стимуляции С и L сегментов. Измеренное нами расстояние между С и L раздражаемыми участками составило 5-14 мм (в среднем. 9.84±-1.44 мм; n = 55). Расстояние между местом введения микроэлектрода в мозг и раздражаемыми шейными сегментами измерялось 3.0-6.9 мм (в среднем. 4.63±0.7 мм; n = 211). Вычисленная вышеотмеченным способом скорость проведения для L нейронов составила 8.0-50 м/с (в среднем 24.47±11.32м/с; n = 100). Однако определение скорости проведения для тех же самих нейронов при использовании отношения d2/t2 показало еe снижение. Впервые на наличие разницы в результатах измерения скорости проведения при использовании отношения (d2-d1)/(t2-t1) и d2/t2 обратил внимание Ито с соавт. [9]. Было показано, что при исследовании вестибулоспинальных нейронов кошки второй способ вычисления занижает скорость проведения на 30%. Это обусловлено следующими тремя факторами: 1) наличием скрытого периода возникновения распространяющегося возбуждения под раздражающими электродами [10]; 2) замедлением скорости проведения при вхождении нервного импульса в сому нейрона из-за падения фактора надeжности проведения на участке границы начального сегмента и сома-дендритной мембраны [11-13]; 3) удлинением пути прохождения нервного импульса из-за отклонения исследуемых трактовых волокон [9]. Наше исследование показало, что у лягушки использование второго способа вычисления (d2/t2) занижает скорость проведения на 78%. Важно отметить, что согласно данным Магерини и соавт. [14] и Гринелл [15] в спинном мозге лягушки часто наблюдается отсутствие антидромной инвазии мотонейронов на стимуляцию переднeго корешка. Это может быть обусловленно тем, что область начального сегмента лягушки имеет более высокое для генерации спайка сопротивление, чем таковoe у мотонейронов кошки, что несомненно может задерживать скорость распространения импульса [16]. С учeтом отмеченных факторов скорость проведения для С нейронов (d1/t1) была определена равной 3.4-13.3 м/с (в среднем 7.44±2.07м/с; n = 96). Общая скорость проведения для всех исследованных С и L нейронов составила 3.4-50 м/с (в среднем 16.12±11.83 м/с; n = 196). На pис.2 представлена гистограмма распределения скоростей для С и L нейронов.
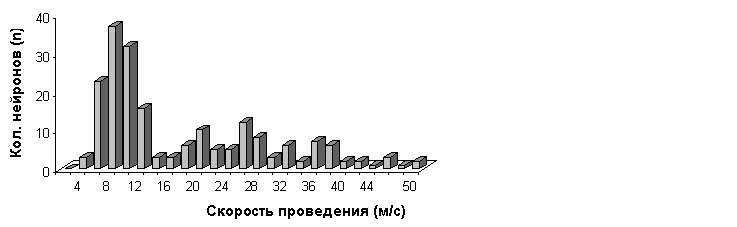 |
лягушки. По оси абсцисс - скорости проведения , в м/с; по оси ординат - количество исследованных
нейронов (n).
Авторы выражают благодарность NFSAT за
содействие в приобретении установки "APP-1 all Purpose Pipette Puller", изготовленные с помощью которой микроэлектроды использовались в проведении
настоящего исследования.
Институт физиологии им. Л.А. Орбели НАН РА
1. Pompeiano O., Brodal A. -
Arch. ital. Biol. 1957 . V. 95 P. 166-195.
2. Бродал А., Вальберг Ф.,
Помпеано О. Вестибулярные ядра. Л. Наука. 1966.
3. Petras J. M. - Brain Res. 1967. V. 6 P.275-324.
4.
Brown L. T. - J. Comp. Neurol. 1974. V. 154 P.
169-188.
5. Шаповалов А. И. Нейроны и синапсы супраспинальных моторных систем. Л. Наука.
1975.
6. Ten Donkellar H. J. In: The Central Nervous System of Vertebrates. Eds. R.
Nienwenhuys, H. J. Ten Donkellar, C. Nicholson. Springer. 1997. V.2 P.
1151-1314.
7. Фанарджян В. В.,
Манвелян Л. Р., Насоян А. М. ДНАН Армении. 2000. T. 100. C.
67-75.
8. Fanardjian V. V.,
Manvelyan L. R., Zakarian V. L., Pogossian V. I., Nasoyan A. M. - Neuroscience. 1999. V. 94. P. 845-857.
9. Ito M., Hongo T., Yoshida M., Okada Y., Obata K. -
Jap. J. Physiol. 1964. V. 14. P. 638-658.
10. Беритов И. С. - Общая физиология мышечной и нервной систем. М. Медгиз.
1959.
11. Brock L. G., Coombs
J. S., Eccles J. C. - J. Physiol. (London). 1952. V. 117. P. 431-
460.
12. Coombs J. S., Curtis
D. R., Eccles J. C. - J. Physiol. (London). 1957. V. 139. P. 232-
249.
13. Экклс Дж. Физиология синапсов. М. Мир. 1966.
14. Magherini P. C., Precht W., Schwindt P. C. - Neurophysiol. 1976. V. 39. P. 459-473.
15. Grinnell A. D. - J. Physiol. (London).
1966. V. 182. P. 612-648.
16. Shwindt P. C. In: Frog Neurobiology, A Handbook. Llinas
Eds. R., Precht W., Berlin, Springer Verlag, 1976. P. 750-764.