ФИЗИОЛОГИЯ
УДК 612.73+612.468
К. В. Казарян, В. Ц. Ванцян, А. С. Тираян, Р. Р. Акопян
Координация быстрой и медленной пейсмекерной
активности
околопузырной зоны мочеточника
(Представлено чл. кор. НАН РА Л. Р. Манвеляном 11/XII 2003)
В ранних исследованиях показано, что каждая
клетка мочеточника может генерировать потенциал действия в ответ на
деполяризацию [1]. Поэтому любая область органа в определенных условиях может
функционировать как пейсмекер. Действительно, последующее изучение
электрофизиологических свойств отдельных областей мочеточника кошек, крыс и
морских свинок наряду с основным околопочечным ритмоводителем выявило
присутствие других автономных пейсмекеров [2-5]. При этом, если центральная
часть мочеточника характеризуется наличием латентных пейсмекеров, способных
активироваться в основном при определенных условиях, то в околопузырной области
собственный спайковый автоматизм, как правило, может наблюдаться и при
нормальных условиях [4]. Последующее изучение области соустья соединения
мочеточника с мочевым пузырем выявило ритмогенез в виде медленных колебаний
мембранного потенциала.
В отличие от колебательного процесса верхнего
пейсмекера (пиелоуретеральное соустье), синхронно создающего распространяющиеся
спайки вдоль мочеточника, медленноволновая активность околопузырной зоны не
координирована со спайковым автоматизмом этой же области. Изучению данного
вопроса и посвящена настоящая работа.
Опыты проводили на 15 взрослых кошках (3-4
кг), наркотизированных нембуталом (50-55 мг/кг) внутрибрюшинно. Описание данной
методики представлено в [3, 6]. Активность околопузырной зоны регистрировалась
введением шарикового электрода в соустье соединения мочеточника с мочевым
пузырем либо перемещением биполярного электрода непосредственно к околопузырной
зоне. Приведенные записи отдельных экспериментов представляют собой данные
регистрации на 7-8 животных.
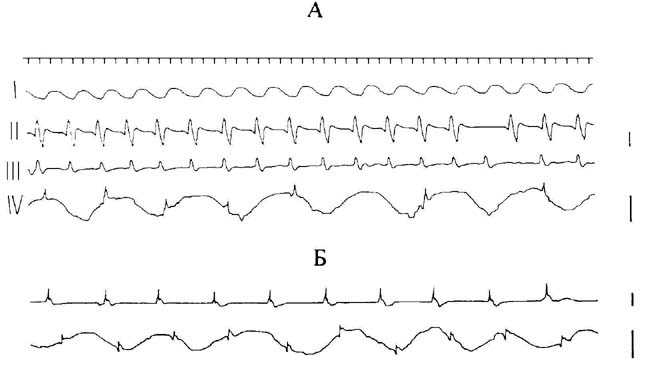
На рис.1,А представлена типичная картина
распространяющейся из области пиелоуретерального соустья вдоль мочеточника
спонтанной активности. Как видно из рисунка, на основе медленных волн синхронно
возникают потенциалы действия, проходящие до мочевого пузыря (первая, вторая и
третья кривые). Введение же электрода непосредственно в область соединения
мочеточника с мочевым пузырем позволило зарегистрировать колебательный процесс
(четвертая кривая), аналогичный верхнему пейсмекеру. Частота выявленных
флюктуаций мембранного потенциала была более чем вдвое реже ритмики
околопочечного автоматизма и несколько уступала частоте спайковой активности
этого же участка [3, 7]. На четвертой кривой рис.1,А представлены
распространяющиеся спайки до области регистрации медленных волн (соустье
соединения мочеточника с пузырем). Никакой координированной деятельности между
верхним и нижним пейсмекерами не наблюдалось, поскольку спайки накладывались на
разные фазы волн. Необходимо отметить, что представленная на рис.1,А активность
соответствует эксперименту, при котором отсутствовал собственный спайковый
автоматизм околопузырной зоны.
Рис.1. Спонтанная активность различных областей
мочеточника кошки при нормальных условиях.
А. Распространяющаяся волна
спайковой активности вдоль органа до соустья соединения мочеточника
с мочевым
пузырем. Активность каждой зоны мочеточника представлена соответствующими
кривыми:
сверху вниз - пиелоуретеральное соустье; область, прилегающая к
соустью; средняя область; соустье
соединения мочеточника с пузырем.
Б.
Наложение спайковой активности околопузырной области на медленноволновый
автоматизм этой
же зоны. Активность каждой зоны мочеточника представлена
соответствующими кривыми: верхняя -
область мочеточника, прилегающая к мочевому
пузырю; нижняя - соустье
соединения мочеточника с мочевым пузырем. Калибровка: 2
мВ, 1 с.
В следующей серии экспериментов нами выявлено
взаимоотношение обоих типов автоматизма исследуемой области в норме. Потенциалы
действия, проходящие до области соустья мочевого пузыря (рис.1,Б), и в данном
случае соответствуют разным фазам колебаний подобно картине наложения на эти
медленные волны проходящих из околопочечного пейсмекера электрических разрядов
(рис.1,А, четвертая кривая).
С целью выявления наличия возможной
координации между электрогенезом быстрой и медленной пейсмекерной активности
проводилась полная перерезка мочеточника в средней его части. Результаты
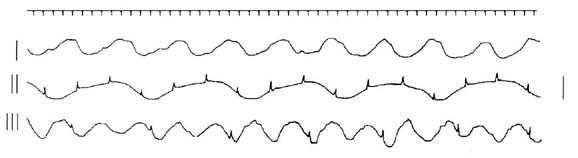
экспериментов в этих условиях представлены на рис. 2. Наблюдались разные типы
взаимоотношения спайкового и медленноволнового автоматизма. Показаны различные
ритмы колебаний, соответствующие разным частотам основного почечного пейсмекера.
Регистрируются колебания как с наложенными на них потенциалами действия, так и
волны без спайков. Kартина активности на первой и третьей кривых рис. 2.
соответствует наблюдениям, когда отсутствуют проходящие спайки либо не каждый из
них достигает области генеза колебаний.
Рис.2. Разные типы медленноволновых колебаний
околопузырной области мочеточника после
перерезки органа. Калибровка: 2 мВ, 1 с.
Вместе с тем при перерезке мочеточника помимо
полной согласованности деятельности спайкового автоматизма и медленных волн в
околопузырной области отмечается также учащение их ритмогенеза.
Известно, что перистальтика гладкомышечной
ткани мочеточника обеспечивается миогенными механизмами [8,9], находящимися под
контролем нейрогуморальной регуляции [6,10]. Возможно, данная регуляция подобно
кишечнику собаки [11] обеспечивает координацию медленной и быстрой пейсмекерной
активности пузырной зоны органа, реализующей свою функциональную роль лишь в
экстремальных условиях.
Институт физиологии им. Л. А. Орбели НАН
РА
Литература
1. Imaizumi Y., Muraki K.,
Takeda M., Watanabe M. - J.Physiol. 1989. V. 411. P. 131-159.
2. Казарян К. В., Тираян А.
С., Маркосян С. А. - Рос. физиол. журн. им. И. М. Сеченова. 1998.
Т. 84 N 5,6. С. 553-555.
3. Казарян К. В., Ванцян В. Ц., Тираян А. С., Акопян Р. Р. - Рос. физиол. журн. им. И. М. Сеченова. 2000. Т. 86. N 12. С.
1656-1661.
4. Казарян К. В.,
Ванцян В. Ц. - Физиол. журн. СССР. им. И. М. Сеченова. 1991. Т.
77. N 10. С. 120-126.
5. Казарян К. В., Ванцян В. Ц., Тираян А. С., Акопян Р. Р. - Рос. физиол. журн. им. И. М. Сеченова. 2001. Т. 87. N 7. С.
953-959.
6. Shuba M. F. - J.Physiol. (L.). 1976. V. 264. P. 853-864.
7. Бакунц С. А. Вопросы физиологии
мочеточников. Л. Наука. 1970.
8. Golenhofen K., Hannappel J. - Pflugers Archiv. 1973. V.
341. P. 257-270.
9. Maggi C.
A., Theodorsson E., Santicioli P., Giuliani S. - Neuroscience.
1992. V. 46. P. 549-559.
10. Santicioli P., Carganico G., Meini S., Guiliani S., Ciachetti A., Maggi
C. A. - Br. J. Pharmacol. 1995. V. 114. P. 1149-1158.
11. Pluja L., Alberti E.,
Fernandez E., Mikkelsen H. B., Thuneberg L., Jimenes M. Am. - J.
Physiol. Gastrointest. Liver Physiol. 2001. V. 281. P. G255-G266.