БИОХИМИЯ
УДК 577.112.612
И. Р. Саакян, Р. Г. Камалян, К. А. Гевондян
Аспартатаминотрансфераза - эффективный регулятор сукцинат
-
зависимого поглощения Ca2+ в митохондриях
сердца и печени
экспериментальных животных
(Представлено академиком А. А. Галояном 28/I 2004)
Аспартаттрансаминаза (АСТ)
играет важную роль в синхронизации энергетического и азотистого обмена,
осуществляемого на уровне митохондрий (МХ). Функционирование фермента связано с
механизмами обмена азотистыми и безазотистыми веществами между митохондриальным
матриксом и цитоплазмой [1]. Конкурируя с цитратсинтазой за оксалоацетат (ОА),
АСТ при обеспечении цикла Кребса ацетил-СоА генерирует ОА, а при дефиците
замыкает цикл, генерируя кетоглутарат (КГЛ). Подобное шунтирование цикла
переаминированием включает и выключает окисление сукцината, генерирующего
восстановительные эквиваленты для синтеза глутамата (ГЛУ). Генерирование НАДН
сукцинатом блокирует вход ацетил-СоА в цикл, способствуя трансаминированию ГЛУ.
Таким образом, осуществляется реципрокное окисление
сукцината, что прослеживается по сукцинатзависимому (СЗ) поглощению Ca2+
МХ и гомогенатами тканей. Шунтирование цикла Кребса переаминированием повышает
вклад в окисление янтарной кислоты [2,3], оказывает сильное регуляторное влияние
на включение в цикл жирных кислот [4] и на использование углеводов в цитозоле.
О. И. Писаренко [5-7] показана взаимосвязь между метаболизмом ГЛУ и аспартата,
синтезом АТФ и стабилизацией работы сердца в условиях его гипоксической
остановки и реперфузии. Cнижение уровня ГЛУ сопряжено с образованием КГЛ и
усилением субстратного фосфорилирования в МХ [8,9].
Сукцинат по интенсивности окисления превосходит НАД-
зависимые субстраты. При его окислении МХ поглощают Ca2+ [10,11]. ГЛУ
и КГЛ уступают ему в этом. Они разнонаправленно воздействуют на окисление
сукцината: ГЛУ стимулирует процесс, по-видимому, за счет устранения ОА, а КГЛ
тормозит [12-15]. Тормозящее действие КГЛ сопряжено с повышением эффективности
фосфорилирования, дыхательного контроля и АДФ/О. Это явление воспроизводится на
МХ и гомогенатах сердца и печени разных животных, прослеживается по СЗ дыханию,
восстановлению НАД и поглощению Ca2+. По-видимому, ограничение КГЛ
окисления сукцината, подобно его активации ГЛУ, может реализоваться через ОА.
Такой подход позволяет по влиянию на окисление сукцината субстратов
переаминирования наблюдать за обратимостью реакции. В качестве чувствительного
теста удобен захват Ca2+ МХ.
В работе исследовали влияние ГЛУ и КГЛ на СЗ поглощение Ca2+
в МХ сердца голубя и гомогенатах сердца и печени крысы.
В работе использовали 20 сизых голубей, из сердец которых
выделяли МХ, а также 30 крыс линии Вистар массой 200 г. Из органов крыс получали
солевые гомогенаты. Для выделения МХ из сердечной мышцы использовали
общепринятую среду (сахароза 300 мкМ, Нереs 10 мМ, ЭДТА 0.5 мМ, рН 7.4) в
соотношении ткань: среда 1 : 10 [16]. Среду суспендирования (без ЭДТА)
использовали в соотношении ткань: среда равном 10 : 1.
Приготовление гомогената сердечной ткани. Сердце после
забоя животного помещали в ледяной раствор следующего состава: 125 мМ КС1, 10 мМ
Hepes, 1 мМ ЭДТА, рН 7.65. Желудочки после иссечения предсердий трижды промывали
вышеуказанной средой, взвешивали и переносили на охлаждаемое льдом часовое
стекло. Cреду гомогенизации обогащали ГЛУ 5 мМ с целью предохранения МХ от
повреждающего влияния продуктов перекисного окисления [13]. Среду добавляли к
сердечной мышце в соотношении 3:1. Тщательно размельченную мышцу растирали (60
с) в неплотном стеклянном гомогенизаторе с помощью тефлонового пестика.
Гомогенат фильтровали через слой капрона, отмечали его конечный объем. Препарат
готов к измерению через 10 мин после забора сердца. Гомогенат (30-40 мг белка/мл)
хранили на льду не более 30 мин. Образцы препарата вносили в среду инкубации по
25 мкл на 1 мл среды.
Приготовление гомогената печени. Процедуры подробно
описаны в [14,17]. После декапитации печень быстро извлекали и помещали в
ледяную среду гомогенизации: 120 мМ КСl, 10 мМ Нepes, рН 7.5. ЭДТА и ГЛУ не
добавляли. Гомогенат фильтровали через двойной слой капрона. Для измерения H+/Ca2+
обмена использовали 200 мкл гомогената с cодержанием белка в 60-70 мг/мл.
Поглощение ионов Ca2+ в митохондриях регистрировали
по противофазному изменению H+ в среде инкубации с помощью
водородного электрода. CaCl2 добавляли порциями по 200 нмолей до
спонтанного выброса [14,15]. Сумма поглощенных катионов характеризует Ca2+
-емкость. По достижении определенного предела возникает спонтанный выброс
накопленного Ca2+. Показано [16,17], что интенсивности входа и выхода
Ca2+ в пробах с и без АДФ различаются. Измерение Ca2+ -емкости
проводили при окислении субстратов с АДФ и без нее [13,15,16]. Оценивали Ca2+
-емкость до и после синтеза АТФ.
Определение скорости синтеза АТФ из АДФ. Измерение
проводили по скорости убыли H+ (защелачиванию среды) после добавления
АДФ.
Инкубационная среда для выделенных МХ сердца голубя
содержала 100 мМ сахарозы, 60 мМ КС1, 1.5 мМ КН2РО4, 1.5
мМ Tris-буфер, рН 7.4; для тканевых гомогенатов сердца и печени крысы - 120 мМ
КСl, 1 мМ КН2РО4, 1 мМ Hepes, рН 7.4, t=25o C.
Субстраты окисления с концентрациями указаны в подписях к рисункам. Везде АДФ
добавляли по 200 мкМ, CaCl2 - по 200 мкМ. МХ и гомогенат вносили в
исследуемую среду (объемом в 2 мл) с заранее добавленным субстратом окисления.
Измерения на тканевых препаратах проводили в течение не более 30-45 мин после их
получения. Во всех исследованиях уровень Ca2+ -емкости и скорость
синтеза АТФ при окислении на сукцинате приняты за 100%.
Белок измеряли методом Лоури [18]. Результаты обрабатывали
по критерию Стюдента и методом парных сравнений (критерий Вилконсона U) [19].
На рис. 1,2 и в табл.1,2 представлены результаты
исследования действия ГЛУ и КГЛ на СЗ накопление Ca2+ выделенными МХ
сердца голубя, а также гомогенатами сердца и печени крысы. Показана
разнонаправленность действия исследованных субстратов: ГЛУ активирует, а КГЛ
тормозит процесс. Выявлено, что КГЛ устраняет вызванную ГЛУ активацию процесса.
С другой стороны, ГЛУ в превышающих концентрациях препятствует этому
блокированию.
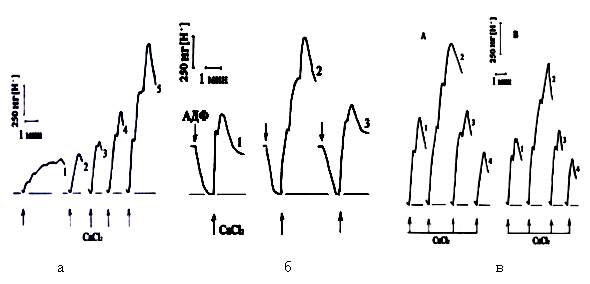
Из рис.1,а видно, что МХ сердца голубя при дыхании на
сукцинате поглощают гораздо больше Ca2+, чем на ГЛУ или КГЛ. СЗ
поглощение Ca2+ усиливается ГЛУ и тормозится КГЛ (см. табл.1).
Примечателен сам характер действия субстратов: ГЛУ (0.6-1мМ) вызывает активацию
окисления во много раз больших количеств сукцината (2.5-5мМ) [11]. Прирост
поглощения Ca2+ варьирует от 96 до 425%, составляя в среднем 294%.
Торможение процесса КГЛ инициируется его относительно большими (5-10 мМ) к
сукцинату концентрациями. При соотношении субстратов 1:2 процесс активируется
незначительно или не изменяется. Активирующее действие ГЛУ в МХ сердца голубя
наблюдается и в условиях их энергизации, т.е. после синтеза АТФ из добавленной
АДФ (рис.1,б). Примечательно, что КГЛ устраняет действие ГЛУ.
Рис.1. Влияние инкубационной среды на поглощение
Ca2+ митохондриями сердца голубя: а) поглощение Ca2+
(100
мкМ) при окислении различных субстратов в МХ сердца голубя. Субстраты: КГЛ 5
мМ(1); ГЛУ 1 мМ(2);
сукцинат 2.5 мМ+ КГЛ 5 мМ(3); сукцинат (4); сукцинат + ГЛУ1
(5). б) Влияние ГЛУ и КГЛ на СЗ синтез АТФ и
поглощение Ca2+ в МХ
сердца голубя. Субстраты:сукцинат 2.5 мМ(1); сукцинат+ГЛУ 1 мМ(2); сукцинат+КГЛ
5
мМ (3). АДФ по 200 мкМ, Ca2+ -100 мкМ. в) cтимуляция ГЛУ (А) или
протимозином (Б) СЗ поглощения Ca2+.
Устранение этой стимуляции КГЛ в
МХ сердца голубя. Субстраты: сукцинат 2.5 мМ (1), сукцинат + ГЛУ 10 мМ +
КГЛ 1
мМ (2), сукцинат + ГЛУ 10 мМ + КГЛ 10 мМ (3), сукцинат + ГЛУ 1 мМ+ КГЛ 10
мМ(4).
Сходные ответы наблюдали на гомогенатах
тканей (см.рис.2,а,б). Торможение реализуется при высоком концентрационном
соотношении КГЛ:ГЛУ, равном 2.5:1 для сердца голубя и 10:1 для сердца и печени
крысы. Торможение в сердце голубя инициируется меньшими дозами КГЛ и в большей
степени, чем в сердце или печени крыс, составляя соответственно 64, 41 и 47% от
исходного (табл.1). Различия во влиянии ГЛУ и КГЛ показаны и на других примерах.
Выявлено, что процедура ресуспендирования осадка МХ сердца голубя средой,
обогащенной ГЛУ, приводит к гораздо большему увеличению поглощения
Ca2+, чем при стандартном методе. Ресуспендирование средой,
обогащенной КГЛ, уменьшает уровень поглощенного Ca2+. В МХ сердца
голубя не только ГЛУ, но и включающий его синтетический пептид активирует СЗ
поглощение Ca2+ (рис.1,в). Действие обоих веществ устраняется КГЛ.
Показана высокая чувствительность СЗ поглощения Ca2+ к изменениям
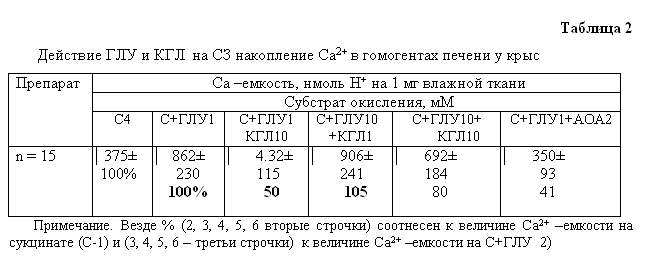
соотношения ГЛУ/КГЛ в гомогенатах печени крысы (рис.2,а,б; табл.2). КГЛ при
количественном преобладании над ГЛУ подавляет процесс. Примечательно, что
показатели синтеза АТФ в сравниваемых пробах практически не изменяются
(рис.2,а).
|
|
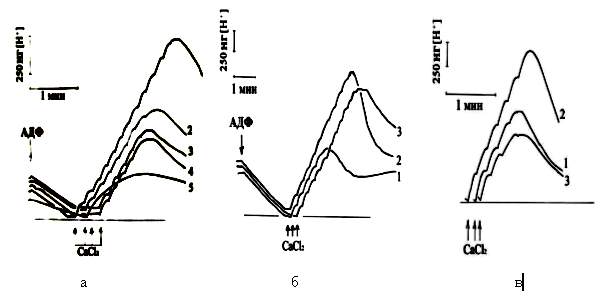
Рис.2. Влияние дыхательных субстратов на СЗ синтез ATP и
поглощение Ca2+ в гомогенатах печени крыс: а)
Влияние ГЛУ и КГЛ на
процессы СЗ синтеза АТФ и поглощения Ca2+. Субстраты: сукцинат
4мМ+ГЛУ 1мМ(1);
сукцинат (2); сукцинат+ГЛУ 1мМ+КГЛ 10 мМ (3); сукцинат + ГЛУ 1
мМ + АОА 2 мМ (4); сукцинат + ГЛУ 1 мМ
+ малонат 2 мМ (5). б) Реципрокность
действия ГЛУ и КГЛ на СЗ поглощение Ca2+. Субстраты: сукцинат 2.5 мМ
+ ГЛУ 1 мМ+КГЛ 10 мМ (1); сукцинат+ГЛУ10мМ+КГЛ1мМ-2;сукцинат+ГЛУ10мМ+КГЛ 10мМ-
3. в) Стимуляция
ГЛУ СЗ поглощения Ca2+. Субстраты: сукцинат 4мМ+ГЛУ
1 мМ (1); сукцинат + ГЛУ 10 мМ + КГЛ 1 мМ (2);
сукцинат + ГЛУ 10 мМ + КГЛ 10 мМ
(3).
По-видимому, действие КГЛ и ГЛУ связано
единым реципрокным механизмом. Взаимодействие между ними осуществляется через
реакцию переаминирования с участием OA и аспарагиновой кислоты. Действительно,
выявлена высокая чувствительность действия КГЛ и ГЛУ к ингибитору трансаминаз,
AOA (рис.2,а, табл.2). АОА снижает активированное ГЛУ поглощение Ca2+
на 59%. При этом показатели синтеза АТФ изменяются лишь незначительно (8-10%).
По сравнению с АОА конкурентный ингибитор активности СДГ - малонат угнетает и
синтез АТФ, и поглощение Ca2+.
Выявлено различие в скорости синтеза АТФ в
солевых гомогенатах сердца крысы, полученных с ЭДТА и без нее. В отсутствие ЭДТА
она уменьшается в ряду субстратов: сукцинат и ГЛУ, сукцинат, ГЛУ и КГЛ, наконец,
сукцинат, ГЛУ и АОА. Добавление АОА приводит не к обычному защелачиванию среды,
а ее закислению. В указанном ряду наблюдаем соответствующее изменение
интенсивности накопления Ca2+. Полученные результаты указывают на
связь переаминирования с фосфорилирующим окислением, которая в определенных
условиях опыта может быть замаскирована.
Таким образом, в настоящей работе показана
(рис.1,2; табл.1,2) взаимосвязь между процессами окисления сукцината, СЗ
накопления Ca2+ и переаминированием ГЛУ. Направление реакции
переаминирования регулируется изменением концентрационных соотношений
участвующих в ней субстратов. Реакция меняет направление под влиянием
возрастающей концентрации какого-либо из названных субстратов, что сказывается
на активации окисления сукцината.
Накопление Ca2+ более эффективно с
сукцинатом, чем с ГЛУ и КГЛ (рис. 1,а, табл.1). ГЛУ усиливает эффект сукцината в
очень низких (0,6 мМ) концентрациях, которые не могут внести вклад в накопление
Ca2+, но достаточны для вовлечения ОА в трансаминазную реакцию,
активации СДГ, что и способствует усилению транспорта Ca2+. Прирост
ГЛУ Ca2+-емкости МХ сердца голубя, отличающихся высокой
чувствительностью к ингибированию ОА, достигает высоких значений (табл.1).
Сходное с ГЛУ действие оказывает ГЛУ-содержащий пептид (рис.1,в).
|
|
Примечательно, что активируемое ГЛУ
накопление Ca2+ устраняется превышающими (10:1) концентрациями КГЛ
(рис.1,а). С другой стороны, в присутствии высоких концентраций ГЛУ тормозящее
действие КГЛ не проявляется. Мы полагаем, что тормозящее действие КГЛ на
окисление сукцината происходит посредством генерации ОА и торможением СДГ. В
зависимости от направления реакции трансаминирования активность СДГ варьирует от
очень высоких до очень низких величин. Накопленный в МХ КГЛ может включаться в
синтез ГТФ в субстратном фосфорилировании, что может привести к замедлению
окисления сукцината. Следовательно, изменение концентрационного соотношения КГЛ
и ГЛУ приводит к активации или торможению окисления сукцината. Это чувствительно
прослеживается по изменению поддерживаемого окислением сукцината накопления
Ca2+ в МХ сердца и печени у различных экспериментальных животных.
Ингибитор трансаминаз АОА предотвращает накопление Ca2+, но на синтез
АТФ не влияет. Малонат тормозит оба исследованных процесса. Торможение
усиливается при совместном применении малоната с арсенитом.
Таким образом, пераминирование ГЛУ и КГЛ
может действовать в качестве реле: включать-выключать поток субстратов в цикл
Кребса, регулировать их пополнение и окисление, обеспечивать адаптационную
пластичность и оптимизировать работу системы в целом. Рассмотренный механизм
может участвовать в осуществлении реципрокной активации окисления сукцината в
МХ. Следовательно, процесс переаминирования может рассматриваться как плавный
переключатель потока субстратов (ди- и трикарбоновых кислот) в цикле Кребса.
Институт хирургии МЗ
РА
Институт биохимии им Г. Х. БунятянаНАН РА
Литература
1. Passarella S., Atlante A.,
Valent D., de Ban L. - Mitochondrion. 2003. V. 2. P. 319-343.
2. Кондрашова М. Н. - Биохимия. 1991. V. 56. P. 388-406.
3. Кондрашова М. Н. - Биофизика. 1989. V.
34. P. 450-458.
4. Randle P. J.
- Circ. Res. 1976. V. 38. P. 108 - 112.
5. Писаренко О. И., Соломатина Е. С., Студнева И. М. -
Биохимия. 1987. V.52. P.543-549.
6. Писаренко О. И., Шульженко В. С., Студнева И. М. -
Кардиология. 2003. N.1. P. 71-75.
7. Писаренко О. И., Студнева И. М., Портной В. Ф. - Бюл. экспер. биол. мед. 1987. T. 54. C. 273-274.
8. Sanborn T., Gavis W.,
Brekowitz S. et al. - Am. J. Physiol. 1979. V. 237. P. H535 - H
541.
9. Takata T., Hiltunen J.
K., Hassinen I. E. - Biochem. J. 1980. V. 192. P. 285-295.
10. Kondrashova M. N.,
Gogvadze V. G., Medvedev B. I., Babsky A. M. - Biochem. Biophys.
Res. Commun. 1982. V.109. P. 376-381.
11. Саакян И. Р. В сб: Терапевтическое действие янтарной
кислоты. Пущино. 1976. C. 201-203.
12. Саакян И. Р. - ДАН АрмССР. 1980. T. 70. N2. C.
110-116.
13. Hansford R. G.,
Hogue B. A., Mildazine V. - J. Bioenerg.Biomembr. 1997 V. 29. P.
89-95.
14. Kondrashova M. N.,
Fedotcheva, N. I., Saakyan I. R., Sirota T. V., Lamasiev K. G., Kulicova M. V.,
Тemnov A. V. - Mitochondrion. 2001. V. 1/3. P.
249-267.
15. Саакян И. Р.,
Карапетян Т. Д., Шердукалова Л. Ф., Пилоян А. Г. А. С. №1455306
СССР. 1988. МКИ С 01 №33/48.
16. Саакян И. Р., Саакян Г. Г. В сб.: Янтарная кислота в
медицине, пищевой промышленности, сельском хозяйстве. ОНТИ ПНЦ РАН. Пущино.
1996. C. 73-186.
17. Caaкян И.
Р., Cаакян С. Г., Kондрашова М. Н. - Биохимия. 2001. T. 66. C.
976-984.
18. Lowry O. H.,
Rosebrough N. J., Farr A. L., Randall R. J. - J. Biol. Chem. 1951.
V 193. P. 265-275.
19. Гублер
Е. В. В кн.: Вычислительные методы анализа в распозновании
патологических процессов. Л. Медицина. 1978.