ФИЗИОЛОГИЯ
УДК 612.826+612.886
Л. П. Манукян
Антидромные и синаптические реакции нейронов ядра Дейтерса
на
стимуляцию верхнего колликулуса
(Представлено чл.-кор. НАН РА Л. Р. Манвеляном 9/III 2004)
Латеральное вестибулярное ядро (ЛВЯ) Дейтерса
и ядро верхнего колликулуса (ЯВК) у млекопитающих являются важнейшими центрами
ствола мозга, контролирующими поведенческие процессы, ориентацию и позу тела в
пространстве [1-4]. В настоящей работе приведены результаты исследования
синаптических и антидромных механизмов вестибуло-тектальных взаимоотношений,
необходимых для завершения серии наблюдений по афферентному контролю активности
вестибуло-спинальных (ВС) нейронов ядра Дейтерса.
Опыты проведены на 68 взрослых кошках массой
2.5-3.5 кг, наркотизированных нембуталом (45-50 мг/кг). Позднее для наркоза
применялся 2 % раствор Рометара или Калипсола (0.5 мг/кг). Биполярными
вольфрамовыми электродами раздражались ипси- (SCi) и контралатеральное (SCc) ЯВК
по следующим стереотаксическим координатам: L 3.3; H 3.3; A 0.1 [5]. ВС нейроны
идентифицировались по их антидромным потенциалам действия (ПД), вызванным
раздражением латерального ВС тракта на уровне С2-3 шейных сегментов спинного мозга. Биполярно
раздражались вестибулярные ядра обеих сторон (0.1-0.15 мА, 0.1-0.5 мс, 1.0-10.0
В). Внутриклеточная активность нейронов ядра Дейтерса регистрировалась
стеклянными микроэлектродами (R=2-10 Мом), заполненными раствором KCl - 2 М.
Точки раздражения и отведения верифицировались гистологически.
Зарегистрирована внутриклеточная активность
280 нейронов ЛВЯ, из них 117 реагировали на раздражение ЯВК.
26 нейронов ЛВЯ отвечали на одиночные
раздражения SCi. Антидромные ПД вызывались в 19 нейронах со скрытым периодом
0.2-1.3 мс (рис. 1, А, B1). Раздражение SCi в нейронах ЛВЯ
вызывало также моно- и полисинаптические
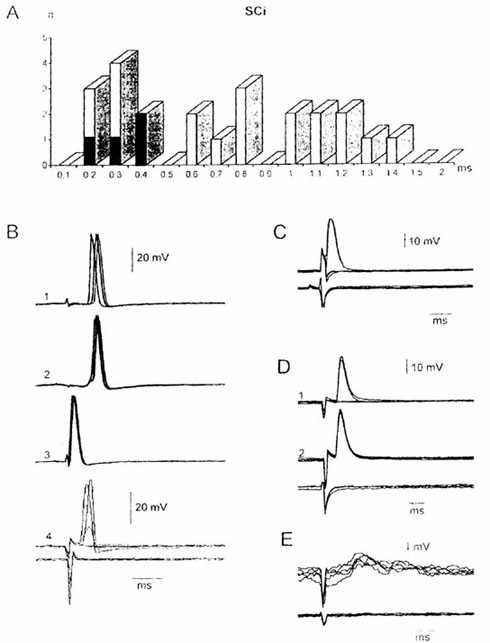
Рис. 1. Антидромные и синаптические реакции нейронов
ядра Дейтерса, вызванные раздражением ипсилатерального ядра верхнего колликулуса
(SCi). А - гистограмма распределения скрытых периодов ПД нейронов ядра Дейтерса
на раздражение SCi. Темные столбики - идентифицированные ВС нейроны. В - ПД
нейрона ядра Дейтерса, вызванные раздражением латерального ВС тракта (1),
ипсилатерального VIII нерва (2), SCi (3) и SCc (4). С - антидромный ПД нейрона
ядра Дейтерса при подпороговом раздражении SCi. D - ортодромный ПД нейрона ядра
Дейтерса, вызванные надпороговым (D1) и пороговым (D2)
раздражением SCi. E - ВПСП нейрона ядра Дейтерса, вызванный стимуляцией SCi.
Здесь и на следующем рисунке представлены потенциалы, зарегистрированные
усилителем постоянного тока при наложении 5-10 пробелов луча; частота повторения
5/с. Нижние записи на В, С, D и E - потенциалы поля при внеклеточном отведении.
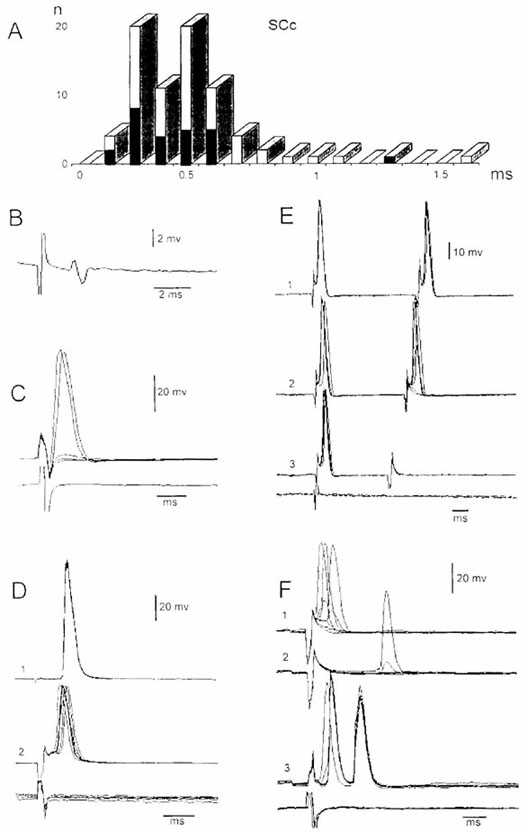
Рис. 2. Антидромные и синаптические реакции ядра
Дейтерса, вызванные раздражением контралатерального ядра верхнего колликулуса
(SCc). A - гистограмма распределения скрытых периодов ответов нейронов ядра
Дейтерса на раздражение SCc. Темные столбики - идентифицированные ВС нейроны. B
- потенциал поля, вызванный раздражением SCc. C - антидромный ПД нейрона ядра
Дейтерса, вызванный пороговым раздражением SCс. D - ПД ВС нейрона ядра Дейтерса,
вызванные раздражением латерального ВС тракта (D1) и SCc
(D2). E - эффекты парной стимуляции SCc с укорочением межстимульного
интервала. F - ПД нейрона ядра Дейтерса, вызванный пороговым (F1) и
подпороговым (F2) раздражением SCc и ипсилатерального VIII нерва
(F3).
ПД и возбуждающие постсинаптические потенциалы (ВПСП)
(рис. 1, D2, E). 5 нейронов из 26 зарегистрированных были
идентифицированны как ВС нейроны (рис. 1, D1, А, темные
столбики).
91 нейрон ядра Дейтерса реагировал на
одиночное раздражение SCc. Антидромные ПД наблюдались в 67 нейронах ЛВЯ. Скрытый
период их возникновения колебался в пределах 0.2-1.1 мс (рис. 2, A). Они
характеризовались постоянным скрытым периодом и возникали по принципу "все или
ничего" (рис. 2, C). 25 из реагирующих на раздражение SCc были
идентифицированы как ВС нейроны (рис. 2, A, темные столбики,
D1). Раздражение SCc в нейронах ЛВЯ вызывало также ортодромные (моно-
(n = 8) и полисинаптические (n = 12)) ПД со скрытым перидом 0.8-8.8 мс (рис. 2,
D2, F1, 2). В одном нейроне ядра Дейтерса раздражение SCc
вызывало моносинаптический ВПСП. Малое количество и слабая выраженность ВПСП в
нейронах ЛВЯ на раздражение ядер верхних колликул, возможно, обусловлены низким
порогом деполяризации для вызова ПД. Наблюдалась конвергенция и дивергенция
влияния со стороны обеих колликул и лабиринтов в нейронах ядра Дейтерса
(рис. 1, В; рис. 2, F). В плане топографии большинство нейронов ядра
Дейтерса, реагирующих на раздражение верхних колликул, распределились
преимущественно в средней и вентральной областях ЛВЯ.
Нейроны ядра Дейтерса посредством своих
восходящих аксонов могут оказывать непосредственное влияние на деятельность ЯВК,
участвуя в процессе координации глазодвигательных реакций [6]. Взаимодействие
между ЯВК и ЛВЯ может осуществляться также через ретикулярную формацию ствола
[2], фастигиального ядра мозжечка [7, 8] и коры мозга [9], с одними и теми же
областями которых связаны обе структуры.
Проведенные эксперименты выявили особенности
взаимодействия между ЯВК и ядром Дейтерса. Последнее, представляя структуру,
основной фукцией которой является нисходящий контроль деятельности спинного
мозга [10,11], проецируется к ЯВК. Стимуляция ЯВК вызывала антидромные ПД в
большинстве нейронов ядра Дейтерса. Последнее свидетельствует о наличии у
аксонов этих нейронов коллатералей, проецирующихся в ЯВК. Таким образом, ВС
нейроны ядра Дейтерса через восходящие коллатерали своих аксонов вовлекают ЯВК в
процесс вестибуло-спинальной интеграции. В свою очередь, наличие
моносинаптических реакций в нейронах ядра Дейтерса при раздражении ЯВК указывает
на модулирующее влияние последних на соматические вестибулярные рефлексы.
Авторы выражают свою благодарность NFSAT за
содействие в приобретении установки "APP-1 all Purpose Pipette Puller". Изготовленные на этой установке микроэлектроды использовались при проведении
настоящего исследования.
Институт физиологии им. Л. А. Орбели
НАН РА
Литература
1. Meredith M. A., Wallace M. T.,
Stein B. E. - Exp. Brain Res. 1992. V. 88. P.
181-186.
2. Sarkisian V. H.
- Arch. ital. Biology. 2000. V. 138. P.
245-353.
3. Sparks D. L.
- Physiol. Rev. 1986. V. 66. P.
118-171.
4. Stein B. E., Meredith
M. A. The Merging of the Senses. MIT Press. Boston. MA.
1993.
5. Berman A. L. The brain stem of the cat. A cytoarchitectonic atlas with stereotaxic
coordinates. Wisconsin. Madison etc. 1968.
6.
Саркисян В. А., Фанарджян В. В. - Сенсорные
системы. 1994. Т. 8. N 1. С. 50-57.
7. Саркисян В. А., Шимон Л. - Физиологический ж. СССР.
1982. Т. 68. С. 768-775.
8. Katoh
Y. Y., Benedek G. - Brain Res. 2003. V. 970. P.
246-249.
9. Sarkisian V. H.,
Fanardjian V. V. - Arch. ital. Biology. 1992. V. 130.
P. 113-126.
10. Fanardjian
V. V., Sarkisian V. H. In: Pompeiano O. and Allum J. H. J.
(Eds). Progress in Brain Research. V. 76. Vestibular Control of Posture and
Locomotion. Elsevier. Amsterdam. New York. 1988. P.
45-60.
11. Wilson V. J., Melvill
Jones G. Mammalian Vestibular Physiology. Plenum Press. New
York. London. 1979.