ФИЗИОЛОГИЯ
УДК 6I2.886:597.82
Академик В. В. Фанарджян, Е. В. Папоян, И. Г. Саркисян, Э.
А. Оганесян
О генезе фокальных потенциалов вестибулярного
ядерного
комплекса лягушки
(Представлено чл.-кор. НАН РА Л.Р. Манвеляном 21/X 2003)
Комплекс вестибулярных ядер представляет
собой центральные структуры, интегрирующие сигналы, поступающие из лабиринта,
мозжечка, спинного мозга, ретикулярной формации, и распределяет свое влияние на
различные двигательные центры [1]. Волокна вестибулярного нерва берут начало в
крупном узле Ganglion vestibularis scarpae, где находится первый нейрон (нейроны
первого порядка) рефлекторной дуги, образуемой вестибулярным нервом.
Периферические отростки нейронов этого узла в составе вестибулярного нерва идут
к лабиринту, т.е. к рецепторам полукружных каналов и преддверья. Центральные
отростки этих нейронов вместе с улитковым нервом образуют VIII пару
черепномозговых нервов, содержащих вестибулярные нейроны второго порядка [2].
Передняя ветвь VIII пары черепномозговых нервов содержит в основном слуховые и
частично вестибулярные волокна.
Цель работы - изучение функциональных
особенностей нейронов II порядка вестибулярного комплекса ядер для выяснения их
роли в формировании целостного рефлекторного акта.
Эксперименты выполнены на перфузируемом
препарате озерной лягушки [3]. Животное анестезировалось 0.1% раствором М5-222
(3-aminobenzoic acid ethyl ester) и при необходимости иммобилизировалось
внутрибрюшинным введением дитилина (0.5 мг/кг). Лягушка перфузировалась
транскардиально ледяным солевым оксигенированным раствором, который содержал (в
мМ) 75 NaCl, 25 NaHCO3, 2 CaCl3, 2 KCl, 0.5
MgCl2, 11 глюкозы. Раствор предварительно насыщали 95% кислорода и 5%
углекислого газа. Скорость протока 1-2 мл/мин. Температура раствора
поддерживалась в среднем на уровне не более 100С, pН 7.4.
Для электрического раздражения использовали
одиночные удары прямоугольного импульса (0.1-0.2 мс; 0.04-0.05 мА), передаваемые
через всасывающие электроды (диаметр 300-350 мкм). Раздражение наносилось с
частотой 0.5 Гц, если ход опыта не меняли. Отводящие электроды, заполненные 3М
KCl, имели сопротивление 3-7 МОм. Регистрация осуществлялась на усилителе
постоянного тока. Применялся компьютерный анализ данных.
Раздражение вестибулярной ветви VIII
черепномозгового нерва в вестибулярном ядерном комплексе ипсилатеральной стороны
вызывало двухкомпонентный фокальный потенциал. Первый короткий компонент
(N0), изредка положительно-отрицательной полярности, в основном
монофазный - отрицательный, более длительный второй (N1) -
отрицательной полярности [4]. Характеристические свойства этих ответов
следующие: N0-компонент регистрировался со скрытым периодом 0.26-0.9
(0.51±0.15) мс, n = 197, фаза восхождения ответа
составляла 0.19-0.05 (0.36±0.09) мс, n = 80, полуспад
0.12-0.16 (0.32±0.11) мс, n = 32, длительность 0.21-1.3
(0.73±0.36) мс, n = 39, амплитуда регистрировалась с
величиной 0.11-3.02 (0.83±0.54) мB, n = 119.
N1-компонент возникал со скрытым периодом 1.0-5.73 (2.16±0.46) мс, n = 172, фаза восхождения составляла 1.1-4.58
(2.99±0.77) мс, n = 142, длительность полуспада -
3.29-15.0 (9.12±4.3) мс, n = 26, амплитуда отрицательной
волны - 0.5-4.64 (1.78±0.78) мB, n = 117. Синаптическая
задержка составляла 1.66±0.3 мс.
N0-компонент, представляющий
быстрое, в основном монофазное отрицательное отклонение, с максимальной
амплитудой регистрировался в латеральных областях ствола мозга [5, 6].
N1-компонент состоял из моносинаптического отрицательного потенциала
(рис. 1).
Рис. 1. Фокальный потенциал вестибулярного комплекса ядер
на раздражение
вестибулярного нерва. Калибровка: 1 мВ; 1 мс.
Для выявления генеза N0- и
N1-компонентов фокального потенциала, регистрируемого в комплексе
вестибулярных ядер на раздражение вестибулярного нерва, применялось тестирование
парным раздражением. Графические данные амплитуды тестируемого ответа при парном
раздражении были соотнесены к интервалам между стимулами. Увеличение амплитуды
тестируемого ответа представляло различие между пиком амплитуды тестируемого
ответа и остаточной деполяризацией от кондиционирующего ответа [7].
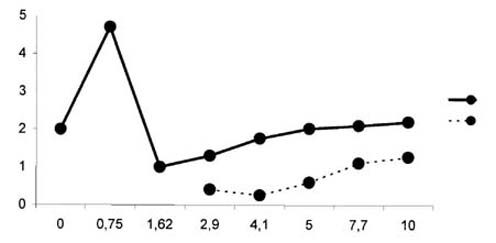
Рис. 2. Влияние парного раздражения на фокальные
потенциалы. Отношение
амплитуды тестируемого N1-компонента потенциала
(ось ординат) к интервалу
между стимулами (ось абсцисс). На 0 отмечено исходное
значение амплитуды
N1-компонента. Сплошная линия - обусловливающий,
пунктирная - тестируемый
стимул.
Регистрацию парного раздражения проводили
наложением регистрируемого процесса и измеряли величину тестируемого ответа на
парную стимуляцию, приложенную с различными интервалами на один и тот же
пресинаптический путь. Величину интервала между стимулами изменяли от
кратчайшего, который все еще позволяет тестируемому ответу начаться после
обуславливающего ответа, до интервала, соответствующего тотальной длительности
обуславливающего ответа, при этом пик N1 волны тест-стимула обычно
равнялся обуславливающему ответу и только иногда мог быть незначительно больше
него. На очень коротких межстимульных интервалах, в среднем на расстоянии 0.75
мс, амплитуда только одного регистрируемого ответа часто превосходила по
величине амплитуду исходного ответа на одиночное раздражение, вероятно,
обнаруживая потенциацию тестируемого ответа. В большинстве случаев ответ на II
стимул появлялся в среднем на pасстоянии между стимулами около 2.5 мс, когда
второй стимул совпадал с вершиной восходящей фазы N1 волны. Если
второй стимул наносился на нисходящую фазу N1-компонента в среднем
при межстимульном интервале 3.5-4.5 мс, ответ тестируемого
N1-компонента уменьшался в амплитуде от 50 до 20 % обуславливающего
ответа (рис. 2).
Таким образом, парное тестирование
свидетельствует о четком различии характеристических свойств N0- и
N1-компонентов фокального потенциала вестибулярного ядерного
комплекса. N0-компонент представляет собой быстрое монофазное
отклонение, отражающее синхронизированную активность афферентного залпа, не
меняется при парном и частотном раздражении или незначительно уменьшается при
коротких межстимульных интервалах, регистрируется с максимальной амплитудой в
поле, где нервные волокна вступают в мозг, представляя пресинаптический
потенциал.
N1-компонент - моносинаптический
отрицательный потенциал подчиняется технике парного раздражения с изменяющимися
межимпульсными расстояниями и представляет собой постсинаптическую деполяризацию
нейронов вестибулярного ядерного комплекса.
Институт физиологии им. Л.А. Орбели
НАН РА
Литература
1. Wilson V.J., Jones G.M.
- Plenum Press. New York, London. 1979. P. 365.
2. Strake H., Biesdorf S.,
Dieringer N. - J. Neurophysiol. 1997. V. 78.
P. 1363-1372.
3. Fanardjian
V. V., Manvelyan L. R., Zakarian V. V., Pogossian V. I., Nasoyan A. M.
- Neurosci. 1999. V. 94. P.
845-857
4. Strake H., Dieringer
N. - J. Neurophysiol. 1999. V. 75. N 5. P.
3087-3101.
5. Фанарджян В.В.,
Манвелян Л. Р., Насоян А. М. - ДАН РАН. 2001. Т. 376. N
4. С. 558-562.
6. Killackey H.P.,
Phoades R.W., Bennet-Clarke C.A. - Trends Neurosci. 1995. V.
18. P. 402-407.
7. Fadiga E.,
Brookhart M. - Neurophysiol. 1962. V. 26. P. 790-805.