
УДК 577.15
Р.М. Сулханян
Сравнительная характеристика действия двух представителей
пролин-богатых гипоталамических нейрогормонов (PRP-1 и PRP-3) в
условиях
интактного животного
(Представлено академиком А.А. Галояном 15/I 2003)
Детальное изучение сигнальных молекул
нейроэндокринной иммунной системы мозга привело к открытию нового семейства
пептидных нейрогормонов (PRP) [1]. Показано, что семейство PRP состоит из 4
пептидов, каждый из которых проявляет специфичные свойства по отношению к
различным функциям организма. Наиболее изучен PRP-1, состоящий из 15
аминокислотных остатков и участвующий в механизмах экспрессии интерлейкинов
(TNF, IL-1, IL-6) в фибробластах, макрофагах и астроцитах. PRP-1 является
иммуномодулятором, активирует образование IL-1, IL-6 и TNF-a в астроцитах и оказывает нейропротекторное действие против
многих токсических продуктов, вырабатываемых в организме и рядом микроорганизмов
[2,3].
Целью данной работы явилось сравнительное
изучение изолированного и комбинированного воздействия PRP-1 и PRP-3 на
активность мотонейронов (МН) спинного мозга (СМ), вызванную на стимуляцию
ипсилатеральных экстензорного (n.gastrocnemius - Gi) и флексорного (n.peroneus
communis - Pi) нервов. В качестве основного критерия сравнительной
характеристики взято действие нейрогормонов PRP-1 и PRP-3 на ранние первичные и
вторичные реакции МН СМ на стимуляцию Gi и Pi.
Эксперименты проводились на 16 зрелых
крысах-самцах (Wistar, 200-250 г). Животные обездвиживались 0.5% дитилином
в дозе 100 мг/кг (в/б) и переводились на искусственное дыхание, затем под
местной анестезией (новокаин) посредством ультразвукового ножа перерезался СМ на
уровнях T2-T3. После фиксации в
стереотаксическом аппарате под новокаиновой анестезией производилась
ламинектомия на уровне L3-L5 в
люмбальной части позвоночника. Стереотаксически дорсо-вентрально ориентированный
стеклянный микроэлектрод (1-2 мкм), заполненный 2М раствором NaCl, вводился в
дорсальные и вентральные рога серого вещества. В сегментах L3-L5 на поврежденной и неповрежденной сторонах СМ
производилась регистрация вызванной активности (ВА) одиночных МН на раздражение
Gi и Pi нервов задних конечностей до и после в/м введения указанных
нейрогормонов в дозе 10g/100г. Регистрация и
последующий анализ проводились на компьютере с помощью специально разработанной
программы по алгоритму, основанному на PETH-гистограммах (Peri-Event Time
Histogram) межспайковых интервалов [2]. Вычислялась постстимульная нейронная
тоническая активность (ТА) [2]. По критерию c-квадрат
оценивалось ее соответствие нормальному, пуассоновскому или равномерному
распределению. Значимость различия фоновой и тонической активности определялась
с помощью двустороннего t-теста Стьюдента. Проводился анализ значимых (P <
0.001) изменений суммарных, усредненных по испытаниям в пределах регистрации,
вызванных импульсных потоков (ИП) до 150 (I150) и
600 (I600) мс на раздражение вышеуказанных нервов, среднего
тонической составляющей - Мср и вклада первичного (ПО) и вторичного раннего
(ВРО) компонентов ВА (I150) в ИП за 600 мс
(I150/I600) при изолированном и сочетанном воздействии
нейрогормонов PRP-1 и PRP-3. Как правило, проводилась антидромная идентификация
МН на его соответствие иннервируемой мышце.
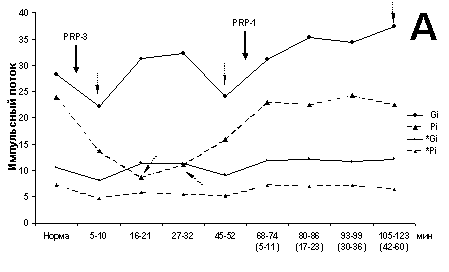
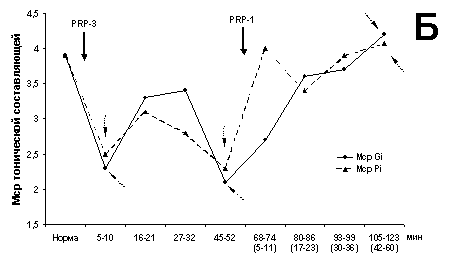
На рис. 1 представлен экстензорный МН
(глубина 1800 мкм), антидромно отвечающий на стимуляцию Gi нерва. Исследовалось
изменение ВА под воздействием PRP-1 с последующим введением PRP-3 на 68 мин. К
25 мин действия PRP-1 почти двукратно возрастает суммарный ИП ВА
(I600), вызванный на раздражение Gi нерва относительно такового в
норме (до введения нейрогормона) с последующим его спадом после введения PRP-3
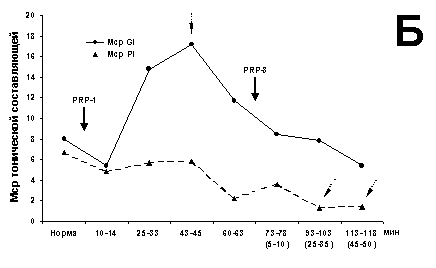
(рис. 1А). Однако среднее значение ТА Мcp изменяется только к 43
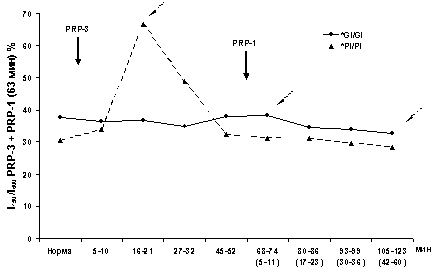
мин (рис. 1Б), тогда как вклад ПО и ВРО достигает минимума на 25 мин и
восстанавливается к 43 мин (рис. 1В), с последующим спадом ИП; он
продолжается и после введения нейрогормона PRP-3, но уже с другой крутизной.
Следует отметить, что этот минимум проявляется за счет ингибиции ВРО ВА. Иными
словами, ТА ВА нейрона на стимуляцию своего нерва свойственно некоторое
характерное поведение. На раздражение Pi максимум потока приходится на 45 мин
(рис. 1А) без значимого изменения Мcp (рис. 1Б).
|
|
Рис. 1. Динамические характеристики влияния PRP-1 и его
комбинации с PRP-3
(10g/100 г для обоих
препаратов, в/м) на экстензорный МН (глубина 1800 мкм).
Здесь и на рис. 2:
А - суммарный ИП в первые 600 мс (I600) и 150 мс
(I150)
регистрации ВА МН на двустороннюю стимуляцию Gi и Pi; Б -
изменения среднего
значения - Mcp ТА; В - развитие во времени
отношения I150/I600. Сплошная стрелка
с названием
нейрогормона указывает на время его применения, пунктирные
стрелки - на значимые
(P < 0.001) отклонения от общего распределения
рассматриваемого параметра.
Кривые, помеченные кружками, соответствуют
значениям параметра при раздражении
Gi в первые 600 (крупные метки) и 150 мс
(мелкие метки), аналогично меченные
треугольником - Pi. На временной оси
диаграмм в скобках - время воздействия
второго гормона.
Введение PRP-3 на 68 мин действия PRP-1 не
влияет существенно на изменение ИП I600 и Мcp ТА при стимуляции Gi нерва,
однако к 25 мин значительно (до 60%) подавляет ВА и Мcp тонической
составляющей на стимуляцию ипсилатерального Pi нерва. Иными словами, ВА нейрона
на раздражение нерва, не представленного в нем, подавляется уже к 25 мин
сочетанного действия нейрогормонов, хотя и наблюдается почти двукратное
увеличение компонентов ПО и ВРО (рис. 1В).
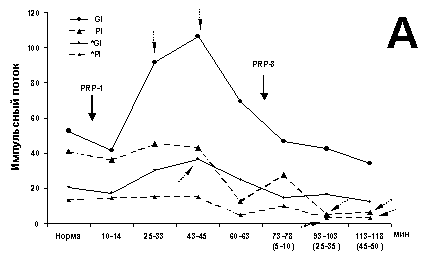
ВА МН (глубина 1600 мкм), также
антидромно отвечающих на стимуляцию экстензорного нерва Gi, но под
предварительным воздействием PRP-3 представлена на рис. 2. Значимое
ингибирующее влияние PRP-3 на ИП ВА (I600) сказывается на 5 и 45 мин
при раздражении своего нерва (Gi) (рис. 2A); аналогично ведет себя среднее
значение тонической составляющей (рис. 2Б). К 16 и 27 мин имеется
незначимый пик ИП и Мcp тонической составляющей между их минимальными
значениями. После введения PRP-1 имеется выраженная тенденция к росту
I600 и Mcp, приводящая к значимому максимуму на 105 мин. К
20 и 32 мин более чем в 2 раза уменьшается поток I600 ВА на
раздражение Pi (рис. 2А). Тенденция к росту Мcp после введения
PRP-1 также приводит к значимому максимуму. К 20 мин двукратно возрастает вклад
ПО и ВРО при раздражении Pi, аналогичный таковому на фоне PRP-3 (рис. 1В).
|
|
Рис. 2. Динамические характеристики влияния PRP-3 и его
комбинации с PRP-1
(10g/100 г для обоих
препаратов, в/м) на экстензорный МН (глубина 1600 мкм).
Таким образом, оба нейрогормона могут
приводить как к активации, так и ингибиции ИП ВА, причем PRP-1 характеризуется
более выраженным возбуждающим, усиливающим импульсацию действием, в то время как
PRP-3 имеет сильно выраженный ингибирующий эффект. Действие обоих нейрогормонов
наиболее выраженно проявляется от 5-10 до 40-60 мин с момента их применения.
Эти особенности ВА проявляются и при их
сочетанном применении. Действие нейрогомонов на ВА оказывается более выраженным
при раздражении МН своим нервом. Оба нейрогормона оказывают существенное влияние
на поздние ответы. Однако PRP-3, по-видимому, характеризуется также определенным
воздействием на тоническую составляющую ИП ВА, понижающим порог эффекторного
ответа. Результаты изучения позволяют предположить, что оба нейрогормона
приводят ВА к некоторому определенному уровню, т.е. к фасилитации нейрона. Тем
не менее наличие определенного взаимовлияния при их сочетанном эффекте не лишает
их самостоятельности.
Институт биохимии им. Г.Х. Бунятяна
НАН РА
1. Galoyan A.A. Biochemistry of novel cardioactive hormones and immunomodulators of the
functional system neurosecretory hypothalamus - Endocrine heart. Moscow. Nauka
Publ. 1997. 240 p.
2. Galoyan
A.A., Sarkissian J.S., Kipriyan T., Sarkisian E.Y., Chavushyan E., Sulkhanyan
R.M., Meliksetyan I.B., Abrahamyan S.S., Grigorian Y., Avetisyan Z.A., Otieva
N.A. - Neurochemical Research. 2001. V. 26. N. 8/9. P.
1023-1038.
3. Galoyan A.A.
- Neurochemical Research. 2001. V.25. N 9/10.
P.1343-1355.