ФИЗИОЛОГИЯ
УДК 611.61.61.617.621
К.В. Казарян, В.Ц. Ванцян, И.Б. Меликсетян, А.С.
Тираян
Воздействие гистамина на активацию латентных пейсмекеров
мочеточника морской свинки
(Представлено академиком В.В. Фанарджяном 3/VII
2002)
Исследование спонтанного ритмогенеза
гладкомышечной ткани мочеточника выявило две исходно активные области,
локализованные в крайних частях органа. В отличиеот околопузырной зоны
(электрогенез отмечен в 60-65% случаев) активность околопочечного пейсмекера
является основным ритмоводителем, обеспечивающим в норме перистательную
деятельность всего органа [1,2]. Средняя же, исходно молчащая, зона мочеточника
характеризуется наличием латентных пейсмекеров, способных активизироваться в
определенных экстремальных условиях [3,4].
Гистамин относится к числу физиологически
активных соединений, специфически влияющих на возбудимость гладкомышечной ткани
мочеточника. В частности, данное соединение способно создавать как медленные
пейсмекерные колебания мембранного потенциала, так и быструю спайковую
активность [5-7].
В литературе на сегодняшний день отсутствует
морфологическая картина локализации исходно активных и молчащих клеточных
образований вдоль мочеточника в отличие от детально исследованной почечной
лоханки [8,9]. Несомненно, определенный интерес вызывает также выявление
изменений морфофункционального состояние клеточных образований при последующей
активации латейных пейсмекеров, в частности, при воздействии гистамина. Это и
послужило целью настоящей работы.
Опыты проводили в условиях in situ на морских
свинках массой 400-600г, наркотизированных нембуталом (50-55 мг/кг). Мочеточники
денервировали путем перерзки корешков чревного и тазового нервов. Спайкове
разряды из околопочечного, среднего и околопузырного участков органа отводили
биполярными электродами. Подробное описание данной методики представленно в
предыдущих работах [4,10]. Ингибирование активности основного ритмоводителя,
локализованного в пиелоуретеральном соустье мочеточника, проводилось путем
охлаждения (приложение льда) данного участка.
Биоэлектрическую активность мочеточника
регистрировали на 8-канальном электроэнцефаллографе (EEG-80, Будапешт).
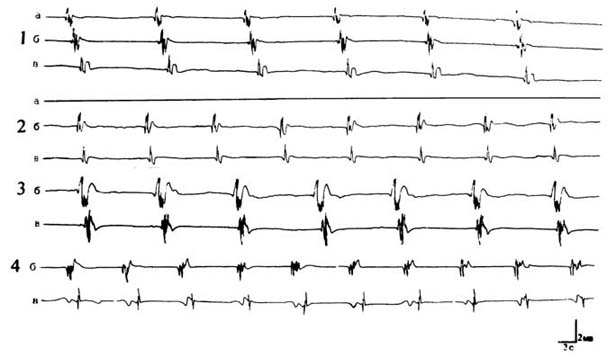
На рис. 1 представлена запись одного из
аналогичных экспериментов, поставленных на 8-10 животных.
Рис. 1. Выявление ингибирования околопочечного пейсмекера
на
спонтанную активность различных областей мочеточника. Активность
каждой зоны
мочеточника представлена соответственно кривыми: а -
пиелоуретеральное соустье,
б - средняя и в - околопузырная области. 1 -
типичная распространяющаяся волна
активности вдоль мочеточника; 2 -
ингибирование околопочечного пейсмекера и
возникновение пейсмекерной
активности средней области и ее распространение в
околопузырную зону;
3,4 - активация латентных пейсмекеров средней зоны под
влиянием
гистамина и распространение октивности в околопузырную зону.
Морфологические исследования
мочеточника проводились после эксперимента гистохимическим методом выявления
ортофосфата [11,12]. Данный метод весьма перспективен не только в
морфологическом аспекте, но и в выяснении функционального состояния самих
структур. Мочеточник фиксировали в 5%-ном растворе нейтрального формалина на
24-48 ч. Из трех отделов мочеточника готовились замороженные срезы толщиной
30-40 мкм, которые переносились в инкубационные смеси, предназначенные для
выявления клеточных ортофосфатов.
На рис. 1, 1 представлена картина
распространяющихся потенциалов действия вдоль всего мочеточника до области,
непосредственно прилегающей к мочевому пузырю. Исследование влияния гистамина
(10-5 -
10-4 моль/л) на исходно молчащую
средную область органа нами приводилось после ингибирования основного
околопочечного ритмоводителя (рис. 1, 2а). Как правило, в 65-70% случаев в
данной области наблюдается возникновение автономного ритмогенеза,
распространяющегося в околопузырную зону (рис. 1, 2, 3). При этом частота
спайков активизированных пейсмекеров несколько превосходит таковую околопочечных
ритмоводителей.
Воздействие гистамина в аналогичных условиях
почти вдвое ускоряет возникновение активности центрального участка мочеточника.
В 30-40% случаев активизированный ритмогенез в этих условиях характеризуется
генерацией потенциалов действия с относительно низкой частотой и более высокими
амплитудами сравнительно с ритмогенезом, создаваемым лишь отсутствием подходящей
волны (рис.1, 3).
|
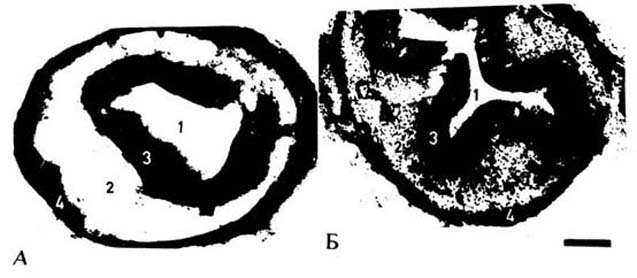
|
Рис. 2. Гистохимическое выявление ортофосфата в клеточных
структурах
средней области мочеточника при воздействии гистамина: А - интактные
морские свнки, Б - после воздействия гистамина; 1 - просвет мочеточника, 2 -
мышечная оболочка, 3 - слизистая оболочка, 4 - адвентициальная оболочка, 5
-
скопление миогенных элементов. Масштаб: 300 мкм.
Гораздо чаще (60-70% случаев) наблюдается
стимуляция активности с частыми и низкоамплитудными спайками (рис. 1, 4). Более
того, как видно из сравнения рис. 1,2 с рис. 1,3 и 1,4, спайки, модулируемые
гистамином, отличаются от нормы также и своей формой.
Морфологическими исследованиями было
подтверждено воздействие гистамина на среднюю зону мочеточника. На рис. 2 для
всех препаратов заметна почти одинаковая степень интенсивности окраски
внутренней - слизистой и наружной - адвентициальной оболочек стенки мочеточника.
Вместе с тем в отношении мышечной оболочки обнаружены заметные различия. Под
влиянием гистамина в средней зоне мочеточника по всей окружности препарата по
сравнению с нормой выявились интенсивно окрашенные миогенные структуры, которые
местами собраны в глыбки (ср. рис. 2,А с рис. 2,Б). Исходя из столь разительной
картины изменения окрашивания данной зоны можно предположить о высоком
функциональном состоянии мышечной оболочки при воздействии гистамина. В этих
условиях отмечается приближение плотности распределения миогенных глыбок в
средней части органа к таковой околопочечной и околопузырной зон в норме, как
это было показано в [1].
Таким образом, при воздействии гистамина
морфофункциональное состояние средней области мочеточника претерпевает
изменения, благодаря которым наблюдается активизация латентных пейсмекеров и
регистрируется ритмогенез, подобно наблюдаемому в иходно активных крайних зонах.
Институт физиологии им. Л.А. Орбели
НАН РА
Литература
1. Казарян К.В., Тираян А.С.,
Маркосян С.А. - Физиол. журн. им И.М. Сеченова. 1991. Т. 77.
N10. С. 120-126.
2. Казарян К.В.,
Ванцян В.Ц., Тираян А.С., Акопян Р.Р. - Росс. физиол. журн.
им. И.М. Сеченова. 2000. Т. 86. N12. С.
1656-1661.
3. Казарян К.В.,
Ванцян В.Ц., Тираян А.С., Акопян Р.Р. - Росс. физиол. журн.
им. И.М. Сеченова. 2001. Т. 87. N7. С.
953-959.
4. Казарян К.В., Ванцян
В.Ц., Меликсетян И.Б., Тираян А.С., Акопян Р.Р. - Росс.
физиол. журн. им. И.М. Сеченова. 2002. Т. 88. N7. С. 925-931.
5. Казарян К.В., Ванцян В.Ц.,
Тираян А.С., Акопян Р.Р. - Росс. физиол. журн. им. И.М.
Сеченова. 1999. Т. 85. N11. С. 1451-1457.
6. Dodel R.C., Hafner D., Borchard U. - Eur. J.
Pharmacol. 1996. V. 318. P. 395-402.
7. Ugaily-Thuoesisus L., Thuiesius O., Angelo-Khattar M., Sabha M.,
Sivanandasinghni P. - Urol. Res. 1988. V. 16. P.
287-293.
8. Gosling Y.A., Dixon
J.S. - Invest. Urol. 1974. V. 11. N5. P.
418-423.
9. Notley R.G.
- B. J. Urol. 1968. V. 40. N1. P.
37-52.
10. Бакунц С.А.
Вопросы физиологии мочеточника. Л. Наука.
1970.
11. Меликсетян И.Б.,
Казарян К.В. В кн.: V конф. молодых физиологов Закавказья.
Баку. 1986. С. 63-64.
12. Чилингарян А.М., Мартиросян Дж.А., Меликсетян И.Б. -
ДАН АрмССР. 1987. Т. 85. N2. С. 83-86.