|
|
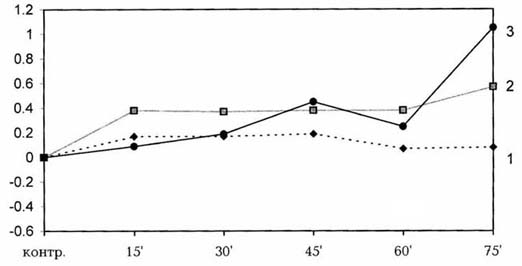
киоторфина интактным крысам. На оси абсцисс - контроль и время
регистрации показателей; на оси ординат - градиент изменения температуры
(DT); 1 - скелетная мышца; 2 - ободочная кишка; 3 - артериальные сосуды.
Эксперименты проводили в следующей последовательности: в первые 30 мин у трех крыс одновременно проводили контрольную регистрацию исследуемых показателей; затем двум экспериментальным крысам внутривенно вводили изобарин (Sigma) в объеме 0.1 мл, содержащий 1 мг чистого препарата на 100 г массы крысы. Третьей крысе вводили физиологический раствор в соответствующем объеме. Далее в течение 40 мин проводили непрерывное термограммирование указанных показателей, после чего всем животным внутривенно вводили киоторфин в дозе 50 мкг/100 r массы тела и термограммирование продолжали в течение 90 мин. Температурные показатели третьей крысы служили контролем для первых двух. На 9 крысах проведено 18 опытов. Результаты экспериментов показали, что киоторфин, введенный на фоне физиологического раствора, индуцировал гипертермию организма (рис.1). Температура ободочной кишки достоверно повышалась и в течение 75 мин доходила до 38.840 С (DT = 0.570). Невысокие колебания температуры скелетных мышц имели фазный характер: в первые 15 мин температура повышалась в среднем на 0.190 С, а к 75-й мин снижалась, доходя до исходного уровня. Температура периферической крови, регистрируемая с кожи хвоста, повышалась на 1.050С (P < 0.01), что свидетельствует о чувствительности b-адренорецепторов кровеносных сосудов к действию препарата.
|
|
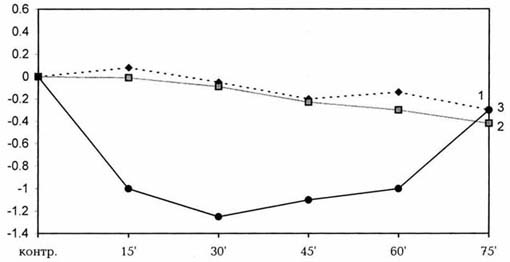
киоторфина на фоне химической десимпатизации. На оси абсцисс -
контроль и время регистрации показателей; на оси ординат - градиент
изменения температуры (DT); 1 - скелетная мышца; 2 - ободочная кишка; 3 -
артериальные сосуды.
Изобариновая десимпатизация приводила к умеренной
гипотермии организма. Наблюдаемые параметры достоверно понижались, составляя
соответственно DT мышц -0.35, ободочной кишки -0.29, артериальных сосудов -0,360С. Киоторфин не только не предотвращал, но и
еще более углублял понижение температуры ''ядра'' крыс, которое регистрировалось
в течение 75 мин и далее. Выявлено, что понижение температурных показателей в
ободочной кишке составило 0.42 (P < 0.01), в мышцах -0.32, а в артериальных сосудах 1.30С (P <
0.01) (рис.2). Под действием киоторфина гипотермия у десимпатизированных крыс
развивалась медленно и только к 75-й мин доходила до достоверных величин. Как у
интактных, так и у десимпатизированных крыс периферические кровеносные сосуды
оказались чувствительнее к киоторфину. Однако колебания температуры крови по
времени были сдвинуты влево и наиболее низкие показатели регистрировались уже
через 15 и 30 мин после инъекции, а к 75-й мин повышались, но не доходили до
исходного уровня. Данный эффект, по-видимому, имел гомеостатическое значение и
был направлен на сохранение температуры ''ядра''. Такое предположение
согласуется с литературными данными [3] о тонизирующем влиянии киоторфина на
кровеносные сосуды. Институт физиологии им. Л. А. Орбели НАН РА
1. Пушкарев Ю. П. , Иванова О. И. В
сб: Симпозиум физиологии пептидов. Л. 1988. С. 145-146.
2. May E. T. , Miller J. - Hormone. Metabol. Res. 1979. V.
11. P. 30-33.
3. Hellone R. F. - J.
Physiol. 1975. V.3. P.29.
4. Попова Н. К. ,
Воронцова И. П. В кн.: Система терморегуляции при адаптации организма к
факторам среды. Новосибирск. 1990. С. 13.
5.
Янский Л. , Выбирал С. и др. В кн.: Нейропептиды и терморегуляция, Минск.
1990. С. 76-90.
6. Брусенцов А. И. -
Рос. физиол. ж. им. Сеченова. 1998. Т.127. N2. С.160-165
7. Данилов Е. И., Графова В. Н. - Бюлл. экспер. биол. и
мед. 1999. Т. 127. С. 160-165.
8. Павлова
И. Ю. , Мационус Э. А. и др - Бюлл. экспер. биол. и мед. 1998. Т. 126.
N7. С. 113-115.
9. Chen P. - J. Med
Chem. 1998. V. 41 (20). P. 3773-3781.
10.
Неbа Н. - Japan. J. Physiol. 1990. V. 79. N 3. P.
263-268.
11. Арутюнян К.Р., Мартиросян С.
Ш. - ДНАН Армении. 2000. Т. 100. N 4. C. 369-372.
12. Авакян О. М. Симпато-адреналовая система. Ереван,
Изд-во АН Арм ССР. 1973. 256 с.