УДК 611.61.61.617.621
Сравнительный анализ ритмогенных свойств
различных
областей мочеточника морской свинки
(Представлено академиком В. В. Фанарджяном 27/VII 2001)
Функциональное значение гладкой мускулатуры
мочеточника заключается в обеспечении необходимой ритмичной перистальтики.
Миогенные механизмы, регулирующие данный процесс, способствуют возникновению
спонтанного электрического ритмогенеза, берущего свое начало в области
пиелоуретерального соустья и распространяющегося вдоль всего органа [1-4].
Вместе с тем в последующих исследованиях электрической спонтанной активности
мочеточника кошки, крысы и морской свинки в зоне, непосредственно граничащей с
мочевым пузырем, был обнаружен отличный по своим показателям от околопочечного
автоматизм [5, 6]. В экспериментах, проведенных на изолированных мочеточниках
морской свинки, помимо двух крайних участков спонтанная активность выявлена
также и в средней области органа, хотя всего в 20% случаев [7]. На целом же
животном при изучении электрофизиологических свойств мочеточника в центральной
его области были обнаружены латентные пейсмекеры, способные становиться
ритмоводителем в определенных экстремальных условиях (нарушение электрической
проводимости, стимуляция физиологически активными веществами).
|
|
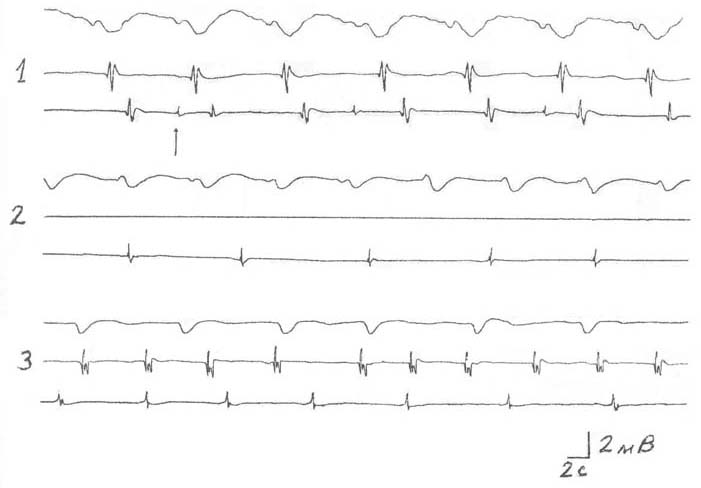
На рисунке, 1 показана описанная типичная активность для мочеточника морской свинки. Ритм распространяющихся спайков, зарегистрированных из средней и околопузырной зон мочеточника, строго соответствует медленным волнам околопочечной области. Перерезка мочеточника между почечной и средней частями приводит к исчезновению распространения потенциалов действия и несколько изменяет картину медленноволновой активности, не изменяя при этом его ритм (рисунок, 2, первая и вторая кривые). Возможно также сохранение и, более того, урегулирование ритмогенеза в околопузырной зоне (третья кривая).
| Эксперимент, гладкие мышцы |
Амплитуда, мВ | Продолжи- тельность, с |
Частота генераций, мин |
Количество осцилляций, шт. |
| Пейсмекерная область |
1.1±0.07
(n 18) |
2.2±0.2
(n 18) |
7.4±0.6
(n 22) |
4.6±0.4
(n 19) |
| Средняя область |
1.35±0.08
(n 22) |
1.98±0.36
(n 16) |
4.5±0.33
(n 23) |
4.1±0.33
(n 24) |
| Околопузырная область |
1.07±0.07
(n 13) |
1.9±0.03
(n 12) |
4.2±0.32
(n 10) |
4.2±0.4
(n 14) |
Через определенный промежуток времени (3-6 мин) более чем в половине из всех наблюдаемых случаев (60-65%) возможно зарождение собственной активности в средней части мочеточника морской свинки (рисунок, 3, вторая кривая). Последующая же перерезка мочеточника между средней и околопузырной областями оказывает заметное влияние на автономный ритмогенез, представленный на третьей кривой (учащается и регулируется автоматизм данной зоны) (рисунок, 3, третья кривая).
| Эксперимент, гладкие мышцы |
Амплитуда, мВ |
Продолжи- тельность, с |
Частота генераций, мин |
Количество осцилляций, шт. |
| Средняя область |
1.26±0.05
(n 10) |
1.8±0.27
(n 11) |
12±0.53
(n 15) |
3.5±0.2
(n 15) |
| Околопузырная область |
0.94±0.1
(n 10) |
1.7±0.26
(n 14) |
6.2±0.68
(n 11) |
4.4±0.56
(n 8) |
Таким образом, подобная постановка эксперимента позволяет
зарегистрировать три различных независимых друг от друга типа активности,
отличных по своим параметрам от исходно наблюдаемой. Институт физиологии им. акад. Л. А.
Орбели НАН РА
1. Бакунц С.
А. Вопросы физиологии мочеточников. Л. Наука.
1970.
2. Kobajashi M, Nagai C.
A., Prosser C. L. - Am J. Physiol. 1969. V. 216. №. 5. P.
1279-1285.
3. Santicioli P.,
Maggi C. A. - Pharmacol Rev. 1998. V. 50. №. 4. P.
683-721.
4. Weis R. M. Im:
Uredynamies (S. Boyarsky ed.). Academic Press. N. Y. 1971. P.
399-410.
5. Казарян К. В.,
Ванцян В. Ц. - Рос. физиол. журн. им. Сеченова. 1991. Т. 77. №. 10. С.
120-126.
6. Казарян К. В.,
Ванцян В. Ц., Тираян А. С., Акопян Р. Р. - ДНАН Армении. 2000. Т. 100. №.
1. С. 88-93.
7. Казарян К. В.,
Тираян А. С., Маркосян С. А. - Рос. физиол. журн. им. Сеченова. 1998. Т.
84. №. 5-6. С. 553-555.
8.
Казарян К. В., Тираян А. С., Акопян Р. Р. - Физиол. журн. СССР. 1990. Т.
76. №. 10. С. 1459-1465.
9. Lang
R. L., Lang J. - J. Urel. 1996. V. 155. P. 332-336.
10. Tahara H. J. - Anat. 1990. V. 170. P. 183-191.