УДК 612.57
Роль киоторфина в регуляции температурного гомеостаза организма
(Представлено академиком В. В. Фанарджяном 6/X 2000)
За последние 10-15 лет выявлен ряд новых
регуляторных пептидов, действие которых отличается широким спектром
[1-3]. Одними из таких пептидов являются киоторфин и неокиоторфин
(тирозин-аргинин), выделенные из мозга быка и организма гибернирующего
длиннохвостового суслика. В работах Павлова и др. [4-7] указано, что
киоторфин участвует в самосогревании гибернирующего организма в период
просыпания. Авторы в течение 90 мин наблюдали повышение ректальной
температуры на 1oС при действии 500 мкг/кг киоторфина, введенного
внутрибрюшинно. Согласно данным других авторов [5-8], дипептид
тирозин-аргинин стимулирует высвобождение метэнкефалина в спинном и головном
мозге, оказывая противосудорожное и антиноцицептивное действие. Однако
малочисленность литературных данных, касающихся функционального спектра
киоторфина, позволяет считать, что изучение его влияния на организм все еще
остается актуальным.
Целью настоящего
исследования явилось сравнение действий разных доз киоторфина на температурный
гомеостаз в условиях нормотермии организма.
Методом многочасового термограммирования в хронических условиях
эксперимента измерялись температурные показатели "ядра" и "оболочки" организма
белых крыс. Измерение температуры "ядра" в области ободочной кишки
проводилось медно-константановой термопарой на глубине 4-5 см. Для
регистрации температуры скелетной мускулатуры "рабочие" спаи термопар с помощью
инъекционной иглы вводились в бедренную мышцу на глубину 1.5-2
см. Измерение температуры артериальных сосудов проводилось с кожи хвоста,
так как хвостовая артерия пролегает под кожей неглубоко. Подобный метод
регистрации данного показателя позволяет судить об участии физических механизмов
в терморегуляции, поскольку непокрытая часть кожи хвоста является своеобразным
теплообменником между организмом мелких животных и средой. "Свободные" спаи
всех термопар помещались в ультратермостат, где поддерживалась эталонная
температура.
Запись исследуемых показателей
проводилась на 12-канальном потенциометре типа ЭПП-09МЗ, который подключался к
выходу фотоэлектрического усилителя марки Ф-116/2. Чувствительность
измерения температуры "ядра" в области скелетных мышц и ободочной кишки была в
пределах 0.013oС, а "оболочки" организма (артериальных сосудов) -
0.13oС. Работа выполнена на 15 крысах, проведено 45 опытов.
Каждый опыт проводился на трех крысах в
следующей последовательности. В первые 30 мин проводилась контрольная
регистрация всех искомых показателей с целью установления плато. Затем двум
экспериментальным крысам в хвостовую вену вводили киоторфин (Sigma, США) в дозе
50 и 100 мкг/100 г массы животного. Препарат перед самым введением
растворяли в физиологическом растворе в объеме 0.1 мл/100 г. Третьей,
контрольной, крысе вводили физиологический раствор в соответствующем
объеме. Регистрация температурных показателей продолжалась в течение 75-90
мин. Все опыты проводились в одно и то же время суток, в хронических
условиях, при температуре окружающей среды в пределах термонейтральной
(22-23oС ). Каждое животное помещалось в специальную камеру (к
которой приучали заранее), ограничивающую его движения, однако исключающую
выраженные эмоциональные реакции. Полученные результаты обрабатывались
методом статистического анализа по критериям Стъюдента.
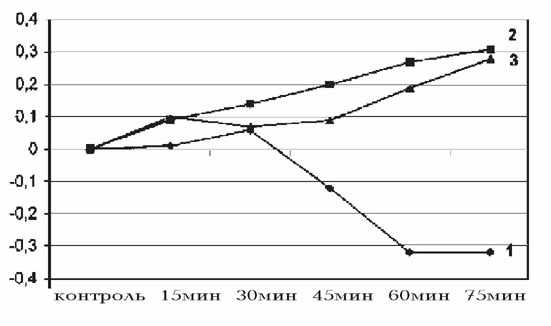
Регистрация исследуемых показателей выявила,
что киоторфин вызывал повышение температуры ободочной кишки уже через 15 мин
после внутривенной инъекции. Гипертермия постепенно усиливалась и достигала
максимума к 75-й мин. Эффект был дозозависимым: введение 100 мкг/100 г
препарата повышало температуру ободочной кишки в среднем на 0.48oС (P
< 0.02), а 50 мкг/100 г - на 0,29oС (P < 0.1; рис. 1).
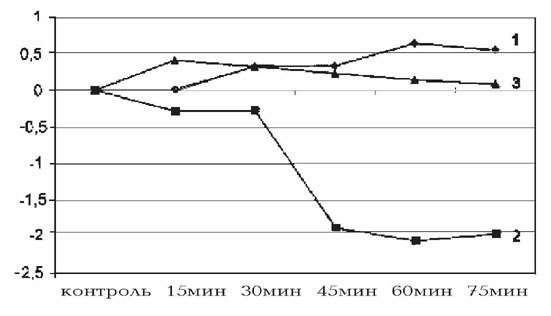
Измерение температуры скелетных мышц выявило
обратный дозозависимый эффект киоторфина. Из рис. 2 видно, что 50 мкг
препарата вызвали большее повышение температуры скелетных мышц, чем 100 мкг/100
г, соответственно \triangle T=0.31 и 0.25oС. Аналогичные
колебания температуры четко прослеживались в каждый момент
регистрации. Подобная тенденция наблюдалась и в динамике изменений
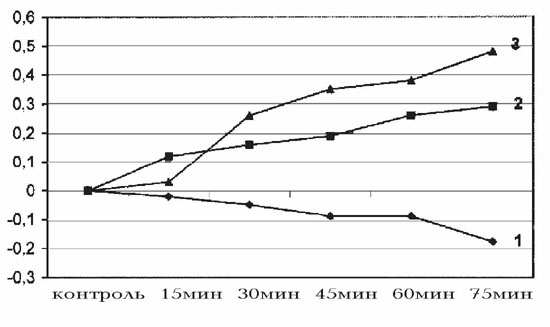
температуры артериальных сосудов. Малые дозы киоторфина вызывали более
сильную вазоконстрикцию (понижение температуры сосудов на 2oС, P <
0.001), чем большая доза препарата, которая вызывала слабую вазодилятацию, с
повышением температуры на 0.5oС (рис. 3).
Температурные показатели контрольных
животных в аналогичных условиях не изменялись либо изменялись
недостоверно. Последнее, вероятно, является следствием гиподинамичного
состояния животных.
|
|
Рис. 1. Действие разных доз киоторфина
на изменение температуры ободочной
кишки в контроле (1), при дозе 50 мкг/100 г (2) и 100 мкг/100 г (3) массы крысы.
На оси абсцисс - контроль и время регистрации показателей,
на оси ординат - градиент изменения температуры (\triangle T).
Анализируя полученные результаты,
можно отметить, что киоторфин нарушает нормотермию организма, вызывая его
гипертермию; оказываемое им действие зависит не только от дозы, но и от
функциональных особенностей эффекторных органов, участвующих в поддержании
температурной константы.
Механизм действия
киоторфина сложен. Сопоставляя полученные нами результаты с данными других
авторов, можно сделать предположение, что киоторфин воздействует опосредованно,
через структуры симпатической нервной системы [9]. Усиливая выброс
норадреналина из симпатических нервных окончаний, он дозозависимо усиливает
термогенез в организме, что прослеживается в ходе регистрации температуры
ободочной кишки. Однако, с другой стороны, киоторфин уменьшает судорожную
активность организма [10], в связи с чем его большие дозы начинают подавлять
сократительный термогенез в мышцах, в то время как малые дозы, наоборот,
стимулируют теплопродукцию в них. Тепло в организме крыс кумулируется также
благодаря вазоконстрикции артериальных сосудов и уменьшению теплоотдачи,
индуцируемому малыми дозами киоторфина. Большие же дозы (100 мкг) вызывают
обратный эффект. По-видимому, киоторфин в малых дозах способен возбуждать
альфа-адренорецепторы симпатической нервной системы (либо непосредственно, либо
через выброс норадреналина), а его большие дозы возбуждают бета-адренорецепторы,
что приводит к слабой вазодилятации.
|
|
Рис. 2. Действие разных доз киоторфина на изменение температуры скелетных
мышц в контроле (1), при дозе 50 мкг/100 г (2) и 100 мкг/100 г (3) массы крысы.
На оси абсцисс - контроль и время регистрации показателей,
на оси ординат - градиент изменения температуры (\triangle T).
|
|
Рис. 3. Действие разных доз киоторфина на изменение температуры хвостовой
артерии в контроле (1), при дозе 50 мкг/100 г (2) и 100 мкг/100 г (3) массы крысы.
На оси абсцисс - контроль и время регистрации показателей,
Таким образом, установленное
терморегуляторное действие киоторфина зависит от его дозы, функциональных
особенностей термоэффекторов организма и опосредуется через структуры
симпатической нервной системы.
Институт физиологии им. Л. А. Орбели НАН РА
1. Арутюнян Р. А., Саакова Л. А.,
Саркисян Д. С., Хачатрян Дж. К. - ДАН АрмССР. 1988. Т. 87. № 4. С.
184-188.
2. Янский Л., Выбирал
С. и др. В кн.: Нейропептиды и терморегуляция. Минск, 1990.
С. 9-31.
3. Мурзенок П. П.
В кн.: Нейропептиды и терморегуляция. Минск, 1990. С. 76-90.
4. Павлов И. Ю., Мационус А. Э.
и др. - Бюлл. экспер. биологии и медицины. 1998. Т. 126. № 7. С.
113-115.
5. Брусенцов А. И. -
Рос. физиол. журн. 1998. Т. 84. № 1-2. С. 115-118.
6. Данилов Е. И., Графова В. Н.-
Бюлл. экспер. биологии и медицины. 1999. Т. 127. № 2. C. 160-165.
7. Chen P., Bodor W. - J. Med. Chem. Sep. 24. 1998. V. 41 (20). P. 3773-3781.
8. Ueda H. - Japn.
J. Pharmacol. 1990, Mar. V. 79(3). P. 263-268.
9. Ковальзон В. И. В кн.: Итоги науки и техники. Сер. физиология человека и животных. 1989.
т. 107, № 6. С. 1512-1514.
10. Покровский В. М., Осадчий О. Е. - Рос. физиол. журн. 1992.
Т. 78. № 4. С. 26-30.