УДК 611.61.617.621
Анализ частотных характеристик латентных
пейсмекеров
мочеточника морской свинки
(Представлено академиком В. В. Фанарджяном 14/IX 2000)
Известно, что сократительная активность
гладкомышечной ткани мочеточника, обеспечивающая направленный транспорт мочи из
почки в мочевой пузырь, контролируется миогенным
пейсмекерным ритмогенезом [1-3]. В ранних исследованиях в области
пиелоуретерального соустья выявлена специализированная высокоавтономная зона,
выполняющая роль водителя ритма всего органа [1]. Вместе с тем в
последующих исследованиях электрической спонтанной активности мочеточника кошки,
крысы и морской свинки в околопузырной области показано наличие автономного
ритмогенеза с показателями, отличными от таковых основного ритмоводителя [4,
5]. Более того, изолированные мочеточники морской свинки проявляют
спонтанную активность и в средней области органа (хотя всего в 20% случаев) [5].
В настоящей работе сделана попытка как
выявить условия активации латентных пейсмекеров мочеточника морской свинки, так
и провести анализ их частотных характеристик.
Показано, что в каждой клетке мочеточника морской свинки деполяризующим
стимулом можно вызвать потенциалы действия, распространяющиеся вдоль органа
[6-8]. Исходя из необходимости обеспечения функциональной активности
мочеточника, на наш взгляд, несомненный интерес вызывают условия активации
латентных пейсмекеров на целом животном.
Работа выполнена в условиях in situ на морских свинках массой 300-500 г,
наркотизированных нембуталом (45-50 мг/кг). Путем перерезки корешков
чревного и тазового нервов денервировали мочеточник. Активность
пейсмекерной области мочеточника, а также распространяющиеся спайковые разряды
из среднего и околопузырного участков органа отводили биполярными
электродами. Ингибирование спонтанной активности пиелоуретерального соустья
производили локальным охлаждением исследуемой области.
Спонтанная активность изолированных полосок
мочеточника морской свинки регистрировалась методом "сахарозного
мостика". После изоляции полоски из различных областей мочеточника
выдерживали в растворе Кребса при температуре 36-37oС в течение
одного часа, затем переносили в соответствующие камеры "сахарозного
мостика". Потенциалы действия отводились каломельными электродами.
Приведенные записи отдельных экспериментов
представляют собой данные регистрации на 7-9 животных.
|
|
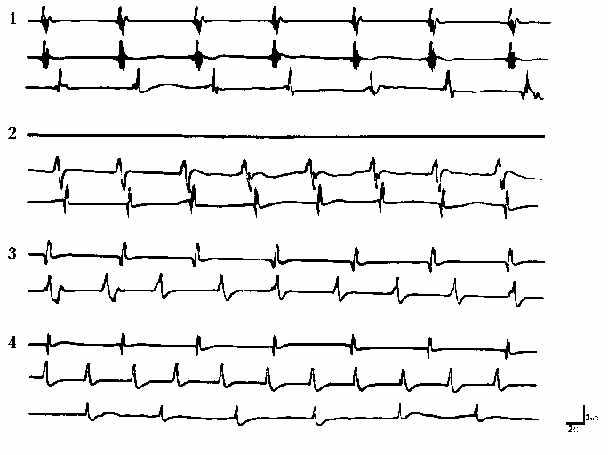
Рис. 1. Влияние ингибирования основного пейсмекера и нарушения
проводимости на активацию латентных пейсмекеров мочеточника морской свинки.
Активность каждой зоны мочеточника представлена соответственно кривыми:
сверху вниз - околопочечная, средняя, околопузырная области.
1 - наличие проходящей волны вдоль мочеточника;
2 - ингибирование активности околопочечного ритмоводителя;
3 - нарушение проводимости между тремя областями;
4 - перерезка мочеточника между тремя областями. Калибровка: 2 с, 2 мВ.
Моторика мочеточника морской свинки по сравнению с кошками и крысами обладает более низким ритмом и колеблется в пределах 7-9 кол/мин [1, 9]. На рис. 1.1 представлена типичная картина спонтанной электрической активности, распространяющейся из области пиелоуретерального соустья до мочевого пузыря. В данной серии экспериментов нами сделана попытка выявить активацию более дистально расположенных областей пиелоуретерального соустья пейсмекеров, не проявляющих спонтанной активности при нормальных условиях. С этой целью исследование спонтанной активности проводилось как при ингибировании основного ритмоводителя, так и при нарушении проводимости вдоль органа.
|
|
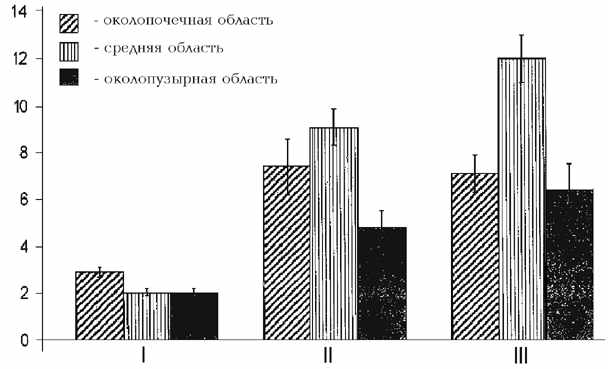
Рис. 2. Характеристики частоты ритмогенеза пейсмекеров средней области
мочеточника морской свинки. I - частота ритмогенеза в условиях "сахарозного
мостика"; II, III - частота ритмогенеза в условиях in situ при нарушении
проводимости и перерезке мочеточника между тремя областями соответственно.
Подавление активности
околопочечного пейсмекера приводит к исчезновению распространяющейся
электрической волны на определенный промежуток времени (7-8 мин). Далее
наблюдается активация молчащих в средней части мочеточника пейсмекеров,
посылающих в область мочевого пузыря электрические импульсы в виде потенциалов
действия (рис. 1.2). Возникающий при указанных условиях электрогенез
характеризуется при этом параметрами, отличными от таковых пиелоуретерального
соустья.
Нарушение проводимости между
крайними областями мочеточника и центральной его частью позволяет наблюдать
возникновение автономных пейсмекерных активностей в каждом из трех участков
органа (рис. 1.3). Последующая перерезка мочеточника в двух частях,
полностью изолирующая друг от друга все три области органа, приводит к изменению
картины активности (рис. 1.4). Изменяются как ритмика пейсмекеров, так
и формы потенциалов действия, в основном, для двух крайних
областей.
Анализ частотных характеристик
автономных пейсмекеров для каждой из рассматриваемых областей в зависимости от
условий их возникновения позволил выявить особенности ритмики латентных
пейсмекеров мочеточника морской свинки (рис. 2). Сравнение параметров
автоматизма пейсмекеров при регистрации спонтанной активности каждой
изолированной области в условиях "сахарозного мостика" с таковыми на целом
животном показало значительное учащение ритмогенеза (ср. рис. 2.I с
2.II и 2.III). В частности, ритмика пейсмекера пиелоуретерального соустья
как при нарушении проводимости (рис. 2.II), так и перерезке мочеточника в
двух частях (рис. 2.III) возрастает более чем вдвое. При этом для всех
трех рассматриваемых условий наблюдается наличие градиента частот между
автоматизмами околопочечного и околопузырного пейсмекеров: преобладает ритм
пиелоуретерального соустья.
Представленные
на гистограмме данные показывают, что в условиях in situ самой высокой частотой
обладают латентные пейсмекеры средней области мочеточника; при полном
разрыве проводящей системы этот показатель увеличивается почти на 30%
(ср. рис. 2.II с 2.III, средние столбики).
Известно, что при нормальных условиях координированная деятельность
пульсации двух крайних областей мочеточника морской свинки определяет полярность
направления перистальтики [9].
Вместе с тем
приведенные в настоящей работе результаты показывают, что центральная часть
мочеточника морской свинки в определенных экстремальных условиях способна
становиться ритмоводителем с высокой частотой. Столь выраженная активация
молчащих пейсмекеров, обеспечиваемая, по-видимому, пластичностью мембранных
свойств, необходима для восстановления функциональной активности органа при
нарушении проводящей системы мочеточника.
Институт физиологии им. Л. А. Орбели НАН РА
1. Бакунц С. А.- Вопросы физиологии мочеточников. Л.: Наука, 1970.
2. Kobayashi M., Nagai T.,
Prosser C. L. - Am. J. Physiol. 1969. V. 216. № 5.
P. 1279-1285.
3. Santicioli
P., Maggi C. A. - Pharmacol. Reviews. 1998. V. 50. № 4.
P. 683-721.
4. Казарян К.
В., Ванцян В. Ц. - Физиол. журн. им. И. М. Сеченова. 1991. Т. 77.
№ 10. С. 120-126.
5. Казарян К.
В., Тираян А. С., Маркосян С. А. - Физиол. журн. им. И. М.
Сеченова. 1998. Т. 84. № 5-6. С. 553-555.
6.
Imaizumi J., Muraki T., Vatanabe M. - J.
Physiol. 1989. V. 411. P. 131-159.
7.
Meini S., Santicioli P., Maggi S. A. - Naunyn-Schmiodeberg's Arch. Pharmacol. 1995. V. 351. P. 79-86.
8. Shuba M. F. - J. Physiol. 1977. V. 264. P. 837-851.
9. Казарян К. В., Ванцян В. Ц.,
Маркосян С. А., Фанарджян С. В. - ДНАН Армении. 1993. 94 № 5.
С. 303-305.