УДК 576.31
Надмолекулярная организация некоторых
биополимеров
миокарда человека
(Представлено академиком В. В. Фанарджяном 24/VIII 2000)
Вопросы надмолекулярной организации ДНП, РНП
и липопротеидов до настоящего времени дискутабельны из-за малых размеров их
элементарных структурных единиц [1, 2]. В настоящей работе изучена in situ
надмолекулярная структура указанных биополимеров нa ультратонких срезах миокарда
больных стенозом левого атрио-вентрикулярного отверстия (ЛАВО) сердца с
использованием трансмиссионной электронной микроскопии и показано их участие в
организации субклеточных структур.
В
хроматине интерфазных ядер мышечных (КМЦ), эндотелиальных и соединительнотканных
клеток миокарда обнаружены кольцевидные частицы с филаментами, одна из которых
длиннее и интенсивнее окрашена, а две боковые, короткие, придают ей некоторую
триангулярность. Длинная филамента у основания несколько
утолщена. Светлые центры кольцевидных структур видны даже в глыбках и
полосе конденсированного хроматина под ядерной мембраной (рис. 1,
2). Их диаметр колеблется в пределах 3-10 нм. Это - ДНП частицы [3,
4]. Мы полагаем, что они имеют сферическую "головку" с филаментами на
уровне экватора и "хвостик" или "ножку", посредством которой прикрепляются к
спирали ДНК (рис. 3, б-1). Ультратонкий срез приходится под разными
углами к осям частиц, поэтому они выглядят по-разному. Вышеописанный вид
характерен для среза, проходящего вдоль длинной оси частицы (рис. 3,
б-2). При избытке контраста она похожа на пирамидную клетку коры головного
мозга (рис. 2, а). Когда срез проходит в плоскости экватора головки -
это окаймленная везикула (рис. 3, б-3), аллегорически названная нами
"аревик" (по-армянски - солнышко) [4]. ДНП частицы остались незамеченными
многими авторами , исследования которых выполнены на разных
органах человека и экспериментальных
животных в норме и
патологии [5-7], что исключает
артефициальность наших данных или
образование описанных частиц только в
условиях патологии.
|
|
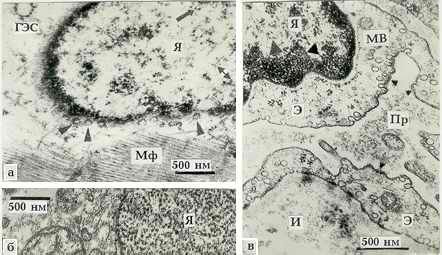
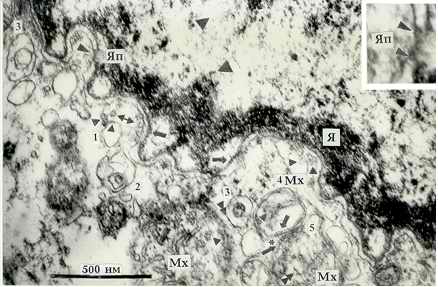
Рис. 1. Фрагмент Я и околоядерной зоны КМЦ левого
желудочка (л. ж.)
сердца больной стенозом ЛАВО Б. Т., 39 лет. ДНП частицы в хроматине
Я отмечены крупными треугольниками. РНП частицы -
равнобедренными
треугольниками (нижняя сторона детали ядерной поры соответствует 14 нм).
В вакуоли (1) маленькими треугольниками отмечены встраивающиеся в
мембрану ВМЧ, а также ВМЧ слившегося с ней пузырька. Стыкующиеся
ВМЧ обозначены сдвоенными треугольниками. Стрелки показывают
поперечно исчеречные участки мембран. Сдвоенная стрелка указывает на
пузырьки, выбухающие из наружной ядерной мембраны и отшнуровавшиеся от
нее. 1-5 - последовательные стадии формирования Мх.
Звездочкой (*) отмечен
намечающийся контакт между Мх 4 и Мх 5, выше он уже сформирован. х 77400.
Условные обозначения:
Я - ядро, ЯП - ядерная пора, Мф - миофибрилла, Мх - митохондрия,
Э - эндотелий, Пр - просвет капилляра, МВ - микропиноцитозные везикулы,
ГЭС - гранулярная эндоплазматическая сеть, И - интерстициальное пространство.
Поскольку химическая композиция ДНП не изменяется при фиксации в глутаровом альдегиде как in situ, так и после его выделения из ядер клеток и подготовке к просмотру в электронном микроскопе [8], можно полагать, что субъединичное строение характерно также и для нативного ДНП. Возможно, его частицы образованы из участков ДНК с повторяющейся последовательностью нуклеотидов, т. е. из так называемых избыточных генов [2], по-видимому, предназначенных для переброски в цитоплазму клетки в тех случаях, когда она нуждается в обновлении цитоплазматических ДНК.
|
|
Рис. 2. а - Фрагмент Я КМЦ л. ж. больной стенозом
ЛАВО Г. Л., 18 лет. РНП
частицы видны по периметру тангенциально срезанных ЯП, отмеченных треу-
гольниками. Стрелка показывает группу ДНП частиц, имеющих форму пирамидных
клеток. Сдвоенная стрелка указывает кольцевидные ДНП частицы. х 30000.
б - Фрагмент Я неидентифированной соединительнотканной клетки миокарда
больной стенозом ЛАВО А. Р., 42 года. Хроматин Я состоит из кольцевидных
субъединиц с филаментами. х 30000.
в - Фрагмент кровеносного капилляра миокарда той же больной; маленькие
треугольники показывают ВМЧ. х 37500.
Молекулярно-биологическими исследованиями также показано образование кольцевидных структур - нуклеосом при различных вариантах денатурации и выделения ДНК из ядер и последующей ренатурации в растворах с различной ионной силой, содержащих ядерные белки - гистоны [8-10], которые являются организаторами суперструктуры, хроматина в виде соленоида, располагаясь в его центре [8, 11]. Пространственное отношение белка к ДНК в ДНП частицах предстоит еще исследовать. Можно только предположить, что начало и конец петли ДНК комплементарно слиты в "ножке", что облегчает их отрыв (называемый в генетике делецией) без нарушения целостности основной нити, а белок покрывает частицу снаружи, формируя ее филаменты.
|
|
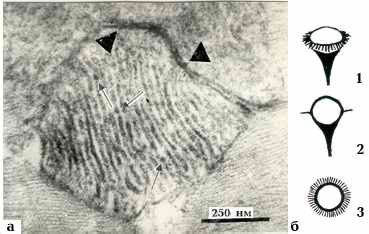
Рис. 3. а - Электронногистохимическая реакция на МВ
в миокарде крысы.
Метод Грахама и Карновского, срок инкубации 1 ч. Локализация осадка в
мембранах крист (стрелки показывают группы ВМЧ с окрашенными
филаментами и "хвостиками", "головки" направлены друг к другу) и оболочках
Мх в зоне межмитохондриального контакта, ограниченного треугольниками.
Снимок из совместной работы Н. Ф. Гусаковой с соавт. [17]. х 98000.
б - Схематическое изображение элементарных частиц биополимеров:
- общий вид, трехмерная модель частиц; 2 - вид на срезе вдоль длинной
оси частицы; 3 - вид сверху, плоскость среза проходит по экватору "головки"
РНП частицы почти такой же
величины и формы. Несколько РНП частиц обнаружено нами в прикрепленной к
мембране гранулярной эндоплазматической сети рибосоме, увеличенной в 410 000 раз
на рисунке из монографии [2], где они почти идентичны с присутствующими здесь
внутримембранными частицами [3, 4], но остались незамеченными авторами или не
комментируются ими. Кольцевидные РНП частицы видны по краям ядерной поры на
профильном срезе (рис. 1, деталь). 8 частиц расположено по ее
периметру на тангенциальном срезе (рис. 2, а). Они известны в
литературе как предшественницы рибосом, синтезированные в ядрышке и
транспортируемые в цитоплазму через ядерные поры [2, 5, 12]. Разница
размеров РНП частиц и рибосом гранулярной эндоплазматической сети весьма
наглядна на рис. 2, а.
Идентичного
строения кольцевидные структуры предположительно липопротеидной природы были
обнаружены в мембранах КМЦ [3, 4]. Они четко видны в крупной Мх
(рис. 1). В процессе предполагаемого биогенеза Мх, последовательные
стадии которого пронумерованы на рис. 1, прослежен механизм образования
оболочек Мх из кольцевидных частиц наружной мембраны ядра КМЦ, обрамляющих
отшнуровывающиеся от нее пузырьки по типу микропиноцитозных, а мембран крист -
из субъединиц пузырьковидных крист разрушенных митохондрий [3, 4, 13],
механизмом обволакивания попавших внутрь будущей Мх, получающей свою ДНК,
по-видимому, из ядра. Считают, что структурные белки мембран крист могут
служить организаторами самосборки мембран в Мх [1, 12]. Внутримембранные
частицы (ВМЧ) в трехмерном изображении выглядят так же, как и нуклеопротеидные
субъединицы. Их диаметр (5-9 нм) соответствует размерам выделенных из
мембран липопротеидных глобул, выявляемых также методом замораживания-скалывания
[2, 12, 14]. Эти ВМЧ идентифицируются в тех участках мембран, которые
срезаны косо и имеют размытый вид (рис. 1, 2в). При стыковке ВМЧ по
принципу "голова-хвост" (рис. 1, Мх 5) боковые филаменты, вероятно,
складываются в сторону хвоста, вместе с головкой, создавая некоторую
неравномерность толщины мембраны (рис. 1, Мх 3), которая исчезает после ее
окончательного формирования, возможно, в результате обратимой пространственной
перегруппировки молекул. Указанную неравномерность толщины, не
комментируемую авторами, мы заметили в новообразующихся в телофазе фрагментах
ядерной оболочки [5]. В двух участках внутренняя ядерная мембрана КМЦ
(рис. 1) утолщена и поперечно исчерчена в результате перпендикулярной к
плоскости мембраны ориентации "хвостиков" частиц, образующих мельчайшие канальцы
по типу "микротоннелей" [6]. Через один из них проходит нить толщиной 3
нм. Поперечно исчерчены также некоторые участки оболочек новообразованных
Мх 4 и Мх 5, что, вероятно, повышает их проницаемость для специфических белков
Мх, синтезируемых на рибосомах цитоплазмы [15]. Мембранные перегородки со
скоплением глобулярных частиц описаны в "септированных межмитохондриальных
соединениях" [16], хотя авторы не указывают, каким структурным элементам мембран
они принадлежат. Такой контакт в процессе формирования из ВМЧ виден между
Мх 4 и Мх 5. "Хвостики" ВМЧ наружного листка оболочки Мх 4 направляются к
"мостикам" в оболочке Мх 5 (*), образовывая септы в
зоне контакта, видимые несколько выше. На рис. 3 из совместной работы
[17] по выявлению активности миоглобинпероксидазы в миокарде крысы
ферментоактивными оказались мембраны оболочек Мх в зоне контакта, а также
оболочки ВМЧ крист, ранее не замеченные, причем осадок диаминобензидинового
коричневого обнаружен именно в филаментах, "хвостиках" и контурах "головок" ВМЧ
при отсутствии окраски сердцевины, что подтверждает предполагаемую нами белковую
природу первых. Из приведенных данных следует, что ламеллярная или
глобулярная организация мембраны обусловлены разной ориентацией "хвостов" ВМЧ в
ее толще, а также обратимой пространственной перегруппировкой
молекул. Следует отметить, что срезанные под разными углами к осям ВМЧ,
имеющие вид колец или овалов с электронноплотным веществом на одном полюсе,
обнаружены нами на электронных микрофотографиях эндотелиальных клеток
кровеносного капилляра миокарда человека [6] и надпочечника кролика [7] в норме.
Таким образом, в результате проведенного
исследования предложена однотипная трехмерная модель надмолекулярной организации
элементарных частиц биополимеров ДНП, РНП и липопротеидов, которая объясняет их
полиморфный вид на срезах в различных плоскостях; показан механизм самосборки
мембран из ВМЧ, ориентации "хвостов" которых в толще мембраны, а также
конформационным изменениям молекул придается ведущая роль в формировании ее
электронномикроскопической картины; продемонстрировано участие ВМЧ различного
генеза в новообразовании Мх. Предложенная схема структурной организации мембраны
удовлетворительно объясняет избирательную проницаемость мембраны.
Данные, полученные на модели патологически
измененного миокарда человека и побудившие к пересмотру некоторых существующих
представлений, конечно же, нуждаются в подтверждении на других моделях и другими
методами исследования.
Институт физиологии им. Л. А. Орбели НАН РА
Институт молекулярной биологии НАН РА
1. Лузиков В. Н. - Регуляция формирования митохондрий. M.: Наука, 1980. 316 с.
2. Робертис Э. де, Новинский В.,
Саэс Ф. - Биология клетки. М.: Мир, 1973. 487 с.
3. Аглинцян Т. С. - Матер. конф. АОЭМ "Электронная микроскопия". Ереван, 1997. С. 5
4. Аглинцян Т. С. - Вестн. хирургии Армении, 1999. № 1-2. С. 140-142.
5. Ченцов Ю. С., Поляков В. Ю. -
Ультраструктура клеточного ядра. М.: Наука, 1974. 175 с.
6. Шахламов В. А. - Капилляры. М.: Медицина, 1971. 200 с.
7. Apkarian R. P. - Scanning. 1997. V. 19. P. 361-367.
8. Thoma F., Koller Th., Klug A.
J.- Cell. Biol. 1979. V. 83. P. 403-427.
9. Martinkina L. F., Yurchenko
V. Yu., Kolesnikov A. A., Vengerov Yu. Yu.- Proc. of 14-th
International Congress on Electron Microscopy, Cancun, Mexiko, 1998,
P. 789.
10. Olins A. I.,
Olins D. E.- Science. 1974. V. 183. P. 4122.
11. Hagmann M.- Science. 1999. V. 285. P. 1200-1203.
12. Лeви А., Сикевиц Ф.
Структура и функции клетки. M.: Мир, 1971, 584 с.
13. Нейфах А. А. В кн.: Митохондрии. Структура и функции. М., 1966. С. 29-42.
14. Структура и функции
биологических мембран. M.: Наука, 1975. 345 с.
15. Schweiger М., Herzog V.,
Neupert V. - J. Cell. Biol. 1987. V. l05. P. 235-246.
16. Бакеева Л. Е., Ченцов Ю. С.
- Митохондриальный ретикулум: строение и некоторые функциональные
свойства. Итоги науки и техники. Общие проблемы биологии. 1989. Т. 9.
103 с.
17. Гусакова Н. Ф.,
Аглинцян Т. С., Кукуртчян Н. С. Матер. конференции АОЭМ
"Электронная микроскопия-97". Ереван, 1997. С. 19.