УДК 577.1527277; 612.8.015
Влияние нового цитокина мозга - галармина на синтез
интерферона-гамма в мононуклеарах человека и на репликацию
вируса энцефаломиокардита в клеточной культуре
(Представлено 6/XII 2000)
Накопленные за последние десятилетия данные
Отдела биохимии нейрогормонов Ин-та биохимии НАН РА им. Г. Х. Бунятяна
свидетельствуют о том, что мозг не только регулирует функции иммунной системы,
но и является одним из ее органов [1, 2]. Основанием для такого утверждения
послужили данные о биосинтезе ряда цитокинов не только клетками макро-
и микроглии мозга, но и нейронами мозга. В частности А. Галоян и сотр.
впервые выделили из нейросекреторных гранул, продуцируемых магноцеллюлярными
ядрами гипоталамуса животных, ряд интерлейкинов и идентифицировали: ИЛ-la,
ИЛ-1б, ИЛ-2, ИЛ-6, ФНОа [3]. Более того, удалось выделить из этих же гранул
совершенно новую семью полипептидов и полностью расшифровать их первичную
химическую структуру [1, 2]. Они обладают выраженными иммуномодулирующими
свойствами. Один из этих полипептидов, содержащий 15 аминокислотных остатков,
названный нами галармином (ГА), оказывает дозозависимое модулирующее действие
на индуцированный ФГА пролиферативный ответ Т-лимфоцитов крови здоровых
доноров. Причем препарат оказывает регулирующее влияние на Т-клеточный
ответ в зависимости от фоновых пролиферативных свойств клеток разных доноров
[4]. Галармин непосредственно воздействует на Т-лимфоциты, что показано
в экспериментах с использованием Jurkat клеток (опухолевые Т-лимфоциты).
Препарат дозозависимо подавляет пролиферацию этих клеток [4]. Галармин
в условиях in vitro разрушает опухолевые клетки мышиных фибробластов (L-929)
[5].
Галармин резко подавляет митотическую активность
этих клеток, меняя также ультраструктуру - фрагментация ядра и цитоплазмы,
разрушение внутренних мембран набухших митохондрий (зрелых L-929 клеток).
Незрелые клетки L-929 под влиянием препарата подвергаются другому виду
изменений в ультраструктуре, появляются пикнотические ядра, ядра с кариорексисом
и кариолизисом, резко выраженный микроплазматоз [5]. Вместе с тем галармин
оказывает влияние на активность каспаз лишь на дифференцированные бутиратом
клетки нейробластомы N2A: в микромолярных концентрациях увеличивает активность
каспаз-2 и каспаз-6, уменьшая активность каспаз-3 и каспаз-9 (основные
протеолитические ферменты, ответственные за гибель клеток [6]). Многочисленные
данные показали иммунокоррегирующие, антибактериальные свойства галармина
[7, 8], а также действие препарата на интерлейкин-2 зависимые функции лимфоцитов
человека в культуре [9]. ГА подавляет ФГА индуцируемую митотическую активность
мононуклеаров.
Вышеприведенные данные свидетельствовали
о возможном действии вновь открытого А. Галояном и сотр. цитокина мозга
- галармина на синтез интерферона-гамма (ИФН-гамма) в мононуклеарах человека
in vitro, а также влияние его на репродукцию вируса энцефаломиокардита
(ВЭМ) в клеточной культуре.
Индукция и титрование ИФН изучены у восьми
здоровых лиц в возрасте от 20 до 40 лет, имеющих нормальный уровень лейкоцитов
и лимфоцитов в крови.
Мононуклеары (МНК) периферической крови
выделяли в градиенте фиколла-верографина и после трехкратного промывания
в среде 199 культивировали в пенициллиновых флаконах (суспензионные культуры)
из расчета 1?106 кл/мл в среде РПМИ-1640 с 10% бычьей сыворотки.
Индукцию ИФН-гамма осуществляли с помощью митогена Т-клеток-ФГА в количестве
10 мкг/мл. Галармин вносили в количестве 40 мкг/мл. Спустя 72 ч с момента
инкубации в термостате клетки осаждали и подсчитывали как в контроле (К),
так и в опытных пробирках (с ФГА, ГА и с ФГА + ГА). Надосадочную жидкость
из опытных флаконов титровали на наличие ИФН биологическим методом. С этой
целью двукратные разведения проб в разведениях с 1:10 до 1:160 вносили
в 96-луночные пластиковые планшеты с монослоем клеток линии НЕр-2 и спустя
18 ч добавляли ВЭМ - 50 тканевых цитопатогенных доз (ТЦД50).
Результаты опытов учитывали спустя 48 ч. За единицу ИФН принимали обратную
величину максимального разведения проб, вызывающего защиту клеток монослоя
от цитопатического действия вируса не менее чем в половине зараженных лунок.
В качестве референс-препарата стандарта для выражения титра ИФН в международных
единицах (ME) использовали рекомбинантный ИФН-a
роферон с активностью 3?106МЕ/мл (Фирма Хоффманн-Ля Рош, Швейцария).
Опыты по изучению действия цитокина на ВЭМ
проводили двумя методами - определение вирулицидности ГА и влияние ГА на
репликацию ВЭМ в клетках НЕр-2.
Для определения вирулицидности две концентрации
ГА (20 и 40 мкг) соединяли с 50 ТЦД50 ВЭМ (равные объемы в среде
Игла с 10% сыворотки) и после инкубации в термостате в течение 2 ч вносили
в лунки планшет с монослоем клеток НЕр-2. В контрольные лунки вносили только
ВЭМ в дозе 50 ТЦД50 после 2-часового выдерживания в термостате.
О вирулицидности препарата судили по степени защиты клеток монослоя от
цитопатического действия вируса, при наличии в контрольных лунках полной
деструкции монослоя.
Влияние препарата на репликацию вируса в
клетках НЕр-2 изучали в различных вариантах опытов: препарат вносили в
клетки за 2 ч до заражения (I вариант), одновременно с заражением (II вариант)
и спустя 2 ч после заражения (III вариант). Контроль - только клетки+ВЭМ.
За антивирусный эффект ГА принимали защиту монослоя не менее чем в половине
зараженных лунок, при условии полной деструкции клеток в контроле вируса.
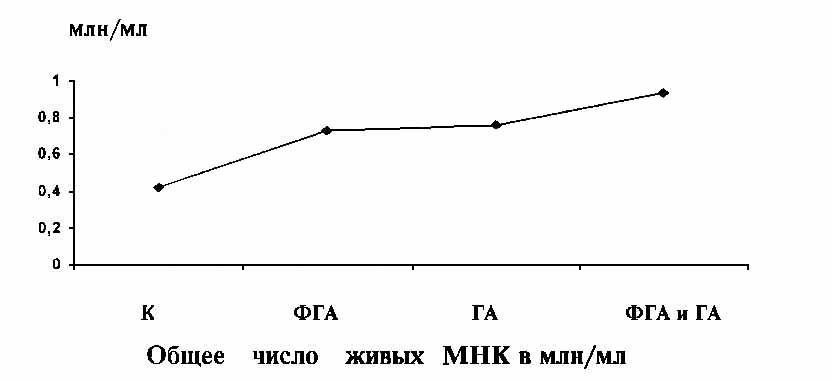
Прежде всего следует отметить, что в суспензионных
культурах МНК галармин в дозе 40 мкг в течение 72 ч инкубации не оказывал
отрицательного действия на жизнеспособность клеток, число которых в тысячах/мл
составляло 765±41. Во флаконах только с ФГА
уровень живых клеток равнялся 730±35. Однако
во флаконах с ФГА и ГА число МНК было наибольшим - 940±51
(разница по сравнению с ФГА статистически достоверна (p < 0.02).
В то же время в контроле (без ФГА) убыль клеток была значительной (422±3.1).
Результаты представлены на рисунке.
Полученные результаты свидетельствуют о
митогенных потенциях ГА, так как уровень живых клеток в течение 72 ч был
почти одинаков при воздействии как ФГА, так и ГА, а их совместное применение
дало достоверный синергидный эффект.
Усредненные результаты титрования проб на
наличие ИФН-гамма представлены в табл. 1. Согласно данным таблицы, ГА не
индуцировал синтеза ИФН (титр < 10 МЕ/мл) ни у одного из доноров. Только
ФГА индуцировал синтез ИФН у всех доноров, усредненный титр составлял 45±6.2.
Но в культурах МНК с ФГА и ГА уровень ИФН был достоверно выше уровня, обеспечиваемого
ФГА (p < 0.02).
| Условия опытов | Титры ГФН в МЕ/мл | Достоверность
p |
| Галармин | < 10 | |
| ФГА | 45±6.2 | |
| ФГА+ГА | 86±9.3 | p < 0.02 |
|
|
|
Влияние нейрогормона на жизнеспособность мононуклеаров |
В предварительных опытах было изучено возможное
токсическое действие ГА на клетки НЕр-2. Наблюдение за клеточной культурой
в течение 5 дней показало, что даже при внесении галармина одновременно
с посевом клеток в дозах 10, 20 и 40 мкг/мл среды он при всех дозах не
влияет на пролиферацию клеток и сроки формирования монослоя по сравнению
с контролем (без галармина).
При изучении влияния ГА на ВЭМ были использованы
дозы - 20 и 40 мкг. Результаты представлены в табл. 2.
|
Условия
опытов |
Варианты
опытов |
Степень защиты клеток НЕр-2
при дозах галармина |
|||
| 20 мкг/мл | 40 мкг/мл | ||||
| Абс. | % | Абс. | % | ||
| Вирулицидность
ГА |
ГА+ВЭМ
(опыт) |
10/20 | 50 | 11/20 | 55 |
| ВЭМ
(контроль) |
0/20 | 0 | 1/20 | 5 | |
| Влияние ГА на
репродукцию ВЭМ |
ГА+ВЭМ
через час |
9/20 | 45 | 11/20 | 55 |
| ГА и ВЭМ
одновременно |
3/20 | 15 | 2/20 | 10 | |
| ГА+ВЭМ
через час |
0/20 | 0 | 1/20 | 5 | |
Примечание: числитель - число сохранивших монослой проб;
знаменатель -
общее число проб; доза вируса - 50 ТЦД50/0.2 мл.
Согласно данным табл. 2, нейрогормон вирулициден,
так как при контакте с ВЭМ вне клеток он нейтрализует цитопатическое действие
вируса. Эффект ГА при обеих дозах почти одинаков. Обе дозы ГА ингибируют
репродукцию ВЭМ в клетках лишь при предварительном внесении препарата в
клетки.
Результаты других вариантов опыта отрицательны,
так как обе дозы ГА защищают монослой от разрушения вирусом лишь в 15-10%
испытуемых проб.
Полученные результаты свидетельствуют о
том, что галармин в концентрации 40 мкг/мл стимулирует пролиферацию лимфоцитов
в коротко живущих культурах мононуклеаров периферической крови здоровых
доноров.
Изученный препарат не индуцирует синтез
ИФН-гамма in vitro, однако он достоверно увеличивает продукцию этого лимфокина
при индукции ФГА. Прайминг эффект ГА можно объяснить пролиферацией лимфоцитов
и активацией их лимфокинной функции. Поскольку ИФН-гамма и ИЛ-2 синтезируются,
в основном, TH1-хелперами, можно предполагать, что ГА преимущественно влияет
на указанную субпопуляцию Т-хелперов.
Для изучения антивирусного действия ГА был
использован вирус энцефаломиокардита мышей, широко распространенный среди
людей и поражающий нейроны и миокард. Вирус размножается в нейронах головного
мозга и с этих позиций является удобной моделью для изучения АВ-активности
ГА.
Вирулицидный эффект препарата реализуется
после контакта его с ВЭМ в течение 2 ч в условиях термостата. Механизм
этого эффекта требует дальнейших исследований, однако можно думать о блокаде
вирусспецифических рецепторов клетки галармином, вследствие чего ВЭМ лишается
способности адсорбцироваться, а следовательно, и проникать в клетки. К
сожалению, о рецепторах пикорнавирусов, к которым относится и ВЭМ, известно
мало, они являются белками, но не выделены, не очищены, а следовательно,
пока не могут быть определены методом ИФА.
Согласно полученным данным, ГА угнетает
репродукцию ВЭМ в клетках НЕр-2 лишь при условии внесения препарата за
1 ч до заражения. Препарат может действовать как на I фазу репродукции
(адсорбция, проникновение и раздевание), так и на II фазу (трансляция и-РНК,
репликация генома и сборка компонентов вируса).
Полученные результаты свидетельствуют о
целесообразности дальнейшего изучения с целью выявления механизмов воздействия
галармина на синтез ИФН и репродукцию вирусов.
Институт биохимии им. Г. Х. Бунятяна НАН
РА
Онкологический научный центр им. В. А. Фанарджяна
Литература
1. Galoyan A. A. Biochemistry
of novel cardioactive hormones and immunomodulators of the functional system
neurosecretory hypothalamus-endocrine heart. Moscow. Nauka publishers,
1997. P. 240.
2. Galoyan A. A. - Neurochem.
Res. 2000. V. 25, N 9. P. 1343-1355.
3. Galoyan A. A., Aprikian V.
S., Markossian K. A., Gurvits B. Ya. - Neurochemistry (RAS & NAS RA).
1998. V. 15. N 4. P. 361-372.
4. Галоян А. А., Шахламов В.
А., Богданова И. М., Малайцев В. В. - Нейрохимия. 2001. T. 17. N 4.
5. Галоян А. А., Шахламов В.
А., Богданова И. Л., Малайцев В. В. - Медицинская наука Армении. 2001.
Т. 41. N 1.
6. Galoyan A. A., Terio N.,
Berg M. J., Marks N. - Neurochemistry (RAS & NAS RA). 2000. V. 17.
N 3. P. 185-188.
7. Априкян B. C., Галоян A.
A. - Медицинская наука Армении. 1999. Т. 39. N 4. C. 29-35.
8. Априкян B. C., Галоян А.
А. - ДНАН Aрмении. 1999. Т. 99. N 4. С. 367-371.
9. Давтян Т. К., Мурадян Е.
Б., Алексанян Ю. Т., Петросян А. А., Галоян А. А. - Нейрохимия. 1998. Т.
15. N 1. С. 45-50.