УДК 597.82:612.886
Электрофизиологические особенности
вестибулоспинальных нейронов лягушки
(Представлено 17/VIII 2000)
Наряду с многочисленными исследованиями мотонейронов
спинного мозга у бесхвостых [1-5], относительно мало работ выполнено по
электрофизиологическому анализу нейронов нисходящих путей спинного мозга
амфибий [6, 7]. Среди последних вестибулярные ядра у бесхвостых занимают
уникальную позицию, поскольку подвержены значительным модификациям, отражающим
изменения в среде обитания (частичный или полный переход на сушу) и развитие
четырехконечностного тела. Уже на этом эволюционном этапе вестибулярные
ядра представляют центральные структуры, оказывающие регулирующее влияние
на двигательные центры. В филогенетическом ряду вестибулярные ядра относятся
к первым супраспинальным группам клеток, которые обособляются от ретикулярной
формации, и их трактовые волокна, нисходящие в спинной мозг, составляют
одну из древних систем надсегментарного контроля [8, 9]. В настоящем сообщении
приведены результаты электрофизиологического изучения особенностей антидромной
активации вестибулоспинальных нейронов на стимуляцию различных уровней
спинного мозга у озерной лягушки (Rana ridibunda).
Эксперименты выполнялись на
перфузированном препарате мозга взрослой лягушки [7]. Животных
глубоко наркотизировали раствором МS-222 (2 мг/кг) или эфиром, охлаждали
колотым льдом и иммобилизировали внутримышечным введением succinylcholina
(1%-ный раствор, 2 мг/кг массы тела, периодически при необходимости). Электрическое
раздражение осуществлялось одиночными прямоугольными ударами тока (0.1-0.2
мс, 0.05-0.4 мА), которые прикладывались через биполярные вольфрамовые
электроды к передней ветви VIII черепного нерва, а также к вентральному
канатику спинного мозга на уровне его шейного (II пара спинномозговых нервов)
и поясничного (VIII-X пары спинномозговых нервов) утолщений с целью антидромной
активации вестибулоспинальных нейронов. Для внутриклеточного отведения
использовались сточенные стеклянные микроэлектроды, заполненные 2 М раствором
хлористого калия с сопротивлением 10-20 МОм. Применялся компьютерный анализ
данных.
Раздражение ипсилатерального вестибулярного
нерва вызывало в вестибулярном ядерном комплексе (ВЯК) фокальные (полевые)
потенциалы, состоящие из двух последовательных негативных компонентов:
N0
(активация вестибулярных афферентных волокон) и N1 (постсинаптическая
активация центральных вестибулярных нейронов). При внутриклеточном отведении
электрической активности вестибулярных нейронов второго порядка на стимуляцию
вестибулярного нерва возникал химически передаваемый моносинаптический
возбуждающий
постсинаптический потенциал, синхронный с
N1
компонентом со скрытым периодом 1.5-2.96 мс.
|
|
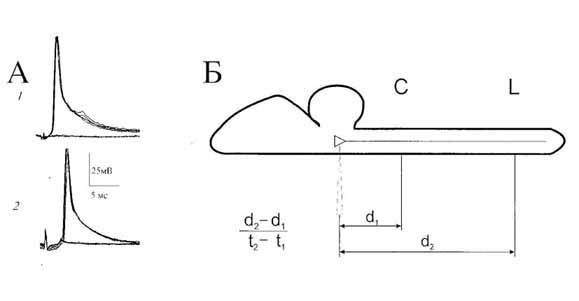
Рис. 1. Антидромная активация вестибулоспинальных
нейронов
на стимуляцию спинного мозга лягушки. А - антидромные
потенциалы
действия С (1) и L (2) нейронов; Б - схема измерения
скорости проведения возбуждения по волокнам вестибулоспинального
тракта;
d1 и d2 - расстояния,
измеренные по вентральной поверхности
спинного и продолговатого мозга от участков раздражения
шейных (С)
и поясничных (L) сегментов до места входа микроэлектрода
в мозг:
t1 и t2 - скрытые
периоды антидромных ответов нейрона на стимуляцию
шейных и поясничных сегментов спинного мозга, соответственно.
В ответ на раздражение вестибулоспинального
тракта в нейронах ВЯК возникали антидромные потенциалы действия, которые
характеризовались при применении различных интенсивностей раздражения коротким
и фиксированным скрытым периодом, короткой рефрактерностью, способностью
воспроизводить высокочастотное раздражение и отсутствием предшествующего
медленного препотенциала. Клетки, которыe могли быть антидромно активированы
только на раздражение шейного отдела спинного мозга, были обозначены C
нейронами. Эта группа клеток включала нейроны, проецирующиеся в шейный,
грудной и верхний поясничный отделы спинного мозга (рис. 1, А 1). Клетки,
которые активировались при раздражении и поясничных сегментов, обозначались
L нейронами. Их аксоны доходили до пояснично-крестцовых отделов спинного
мозга (рис. 1, А 2). Скрытые периоды антидромных потенциалов действия С
нейронов составляли 0.57- 3.6 мс (в среднем 1.57±1.69
мс; n=121). Антидромные потенциалы L нейронов возникали со скрытыми
периодами 1.3-3.89 мс (в среднем 2.18±2.25 мс;
n=94).
Как показано на рис. 1, Б, когда клетка
ВЯК антидромно активируется на стимуляцию шейного (C) и поясничного (L)
отделов спинного мозга, скорость проведения по ее аксону может быть высчитана
как отношение (d2-d1)/(t2-t1),
при котором t1 и t2 являются величинами
скрытых периодов нейрона на раздражение шейного (C) и поясничного (L) сегментов
спинного мозга (рис. 1), а d1 и d2
представляют расстояние между местом введения микроэлектрода в ствол мозга
и участками стимуляции C и L сегментов. Измеренное нами расстояние между
C и L раздражаемыми участками составило 7.0-13.0 мм (в среднем 9.55±
9.66 мм; n=97). Расстояние между местом введения микроэлектрода
в мозг и раздражаемыми шейными сегментами измерялось 3.8-9.9 мм (в среднем
6.22± 6.34 мм; n=125). Вычисленная вышеотмеченным
способом скорость проведения для L нейронов составила 2.5-42.8 м/с (в среднем
15.84±18.42 м/с; n=108). Однако определение
скорости проведения для тех же самых нейронов при использовании отношения
d2/t2
показало ее снижение. Впервые на наличие разницы в результатах измерения
скорости проведения при использовании отношения (d2-d1)/(t2-t1)
и d2/t2 обратил внимание Ито с соавт.
[10]. Было показано, что при исследовании вестибулоспинальных нейронов
кошки второй способ вычисления занижает скорость проведения на 30%. Это
обусловлено следующими тремя факторами: 1) наличием скрытого периода возникновения
распространяющегося возбуждения под раздражающими электродами [11]; 2)
замедлением скорости проведения при вхождении нервного импульса в сому
нейрона из-за падения фактора надежности проведения на участке границы
начального сегмента и сома-дендритной мембраны [12-14]; 3) удлинением пути
прохождения нервного импульса из-за отклонения исследуемых трактовых волокон
[10]. Проведенное нами исследование показало, что у лягушки использование
второго способа вычисления (d2/t2)
занижает скорость проведения на 78%. Важно отметить, что согласно данным
Магерини и соавт. [3] и Гринелл [2] в спинном мозгу лягушки часто наблюдается
отсутствие антидромной инвазии мотонеиронов на стимуляцию переднего корешка.
Это может быть обусловлено тем, что область начального сегмента лягушки
имеет более высокое для генерации спайка сопротивление, чем таковая у мотонейронов
кошки, что несомненно может задерживать скорость распространения импульса
[5]. С учетом отмеченных факторов скорость проведения для С нейронов (d1/t1)
была определена равной 3.6-23.5 м/с (в среднем 10.67±11.54
м/с; n=128). Общая скорость проведения для всех исследованных С
и L нейронов составила 2.5-4.28 м/с (в среднем 13.04±15.085
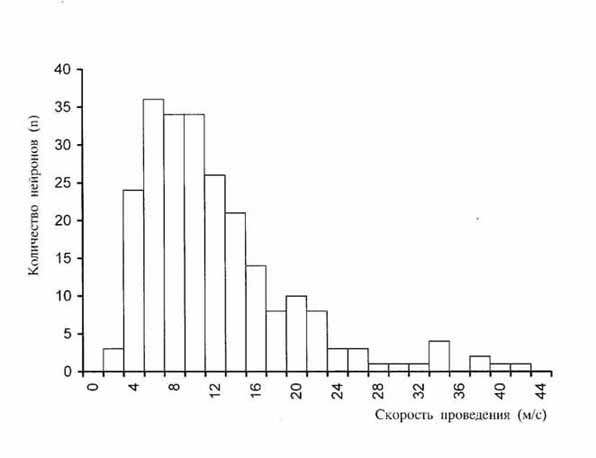
м/с; n=236). На рис. 2 представлена гистограмма распределения скоростей
для С и L нейронов.
|
|
Институт физиологии им. Л. А. Орбели НАН РА
Литература
1. Araki T. - Jap. J. Physiol.
1960. V. 10. P. 518-532.
2. Grinnell A. D. - J. Physiol.
(London). 1966. V. 182. P. 612-648.
3. Magherini P. C., Precht W.,
Schwindt P. C. - 1976. Neurophysiol. V. 39. P. 459-473.
4. Magherini P.C., Precht W.,
Schwindt P. C. - 1976. Neurophysiol. V. 39. P. 474-500.
5. Shwindt P. C. - In: Frog
Neurobiology, A Handbook. Eds. R. Llinas,W. Precht. Berlin, Springer Verlag,
1976. P. 750-764.
6. Precht W., Richter A, Ozawa
S., Shimazu H. - Exp. Brain Res. V. 19. P. 377-393.
7. Fanardjian V. V., Manvelyan
L. R., Zakarian V. L., Pogossian V. I., Nasoyan A. M. - Neuroscience. 1999.
V. 94. P. 845-857.
8. Shapovalov A. A. - Rev. Physiol.
Biochem. Pharmac. 1975. V 72. P 1-54.
9. H. J. Ten Donkellar - In:
The Central Nervous System of Vertebrates. Eds. R. Nieuwenhuys, H. J. Ten
Donkellar. C. Nicholson. Springer. 1997. V 2. P. 1151-1314.
10. lto M., Hongo T., Yoshida
M., Okada Y., Obata K. - Jap. J. Physiol. 1964. V 14. P 638-658.
11. Беритов И. С. - Общая физиология
мышечной и нервной систем. М.: Медгиз, 1959.
12. Brock L. G., Coombs J. S.,
Eccles J. C. - J. Physiol. (London). 1952. V. 117. P. 431-460.
13. Coombs J. S., Curtis D.
R., Eccles J. C. - J. Physiol. (London). 1957. V. 139. P. 232-249.
14. Экклс Дж. - Физиология синапсов.
М.: Мир, 1966.