УДК 612.58+615.94
Действие яда гюрзы на терморегуляторные показатели
организма крыс в зависимости от возраста и пола
(Представлено академиком В.В. Фанарджяном 28/XI 1999)
Змеиные яды обладают широким спектром физиологической
активности и действуют на многие
Ранее нами был изучен гипотермический эффект
яда гюрзы при его внутривенном введении в организм [5], а также его воздействие
на температурный гомеостаз организма, опосредованное симпатической нервной
системой. Данные, полученные в экспериментах с изобариновой десимпатизацией,
привели к выводу, что яд действует через симпатическую нервную систему
и оказывает симпатомиметическое действие, но не адренэргическое, ибо в
этом случае он компенсировал бы изобариновое истощение катехоламинов.
Поскольку активность и лабильность симпатической
системы, а также реакция на яд меняются с возрастом и полом [6,7], был
проведен ряд экспериментов, где исследовалось действие яда гюрзы на терморегуляторные
показатели организма в зависимости от пола и возраста, т. е. у молодых
и зрелых самцов и самок крыс.
Методом длительного термографирования у
интактных крыс, а также у крыс разных половых и возрастных групп, медицированных
ядом гюрзы в термонейтральных условиях (21-22°C)
окружающей среды, определяли температуру "ядра" в области ободочной кишки
и бедренных мышц и температуру "оболочки" в области проксимального отдела
хвостовой артерии. Датчиками для регистрации температуры с исследуемых
участков тела являлись термопары диаметром 100 мкр. Регистрация производилась
при помощи 12-канального самопишущего потенциометра. Последний был подключен
к выходу фотоэлектрического умножителя Ф-116/2 с чувствительностью 0,013°C
для определения температуры "ядра" и 0,13°C
- "оболочки" и термокамеры. "Рабочие" спаи термопар вводили: в мякоть бедренных
мышц на глубину 1,5-2,0 см; через прямую кишку в ободочную на глубину 6-7
см; закрепляли над хвостовой артерией на расстоянии 1,5 см от анального
отверстия. "Свободные" спаи всех термопар помещались в ультратермостат
типа U-10, где поддерживалась эталонная температура.
Яд гюрзы вводили в дозе 50 мкг/100 г массы
тела, при LD50, равном 1,9 мг/кг массы тела, и общем
содержании белка, равном 97,8 %. Яд кавказской гюрзы (Vipera lebetina obtusa)
был получен путем механической дойки змей одним из авторов настоящей статьи.
Яд подвергался сушке в чашках Петри над дессикантом в вакуум-эксикаторе
при остаточном давлении -0,9 атм до его полной
"кристаллизации" и доведении до постоянного веса. Стандартизация яда проводилась
в соответствии с наиболее распространенными требованиями по сертификации
данного продукта [8]. За час до начала эксперимента готовился стандартный
раствор с концентрацией 1 мкг/мкл. Крысам со средней массой 250 г вводилось
в среднем 125 мкг яда. После получасовой регистрации исходного фона в трех
регистрируемых областях (ободочная кишка, бедренная мышца и кожа над хвостовой
артерией) подопытным крысам внутривенно вводился яд гюрзы. Контрольные
крысы получали такую же аликвоту физиологического раствора. Работа выполнена
на 26 крысах, проведено 18 опытов.
Во время экспериментов животные находились
в прозрачных камерах с ограниченным объемом, однако при визуальной оценке
их состояния гиподинамический статус не вызывал девиантного поведения,
не было отмечено полипноэ, саливаци или какой-либо выраженной эмоциональной
реакции.
Эксперименты были проведены на двух группах
крыс. В первой группе были половозрелые крысы обоих полов (в возрасте старше
одного года), во второй - молодые (4 - 5-месячные).
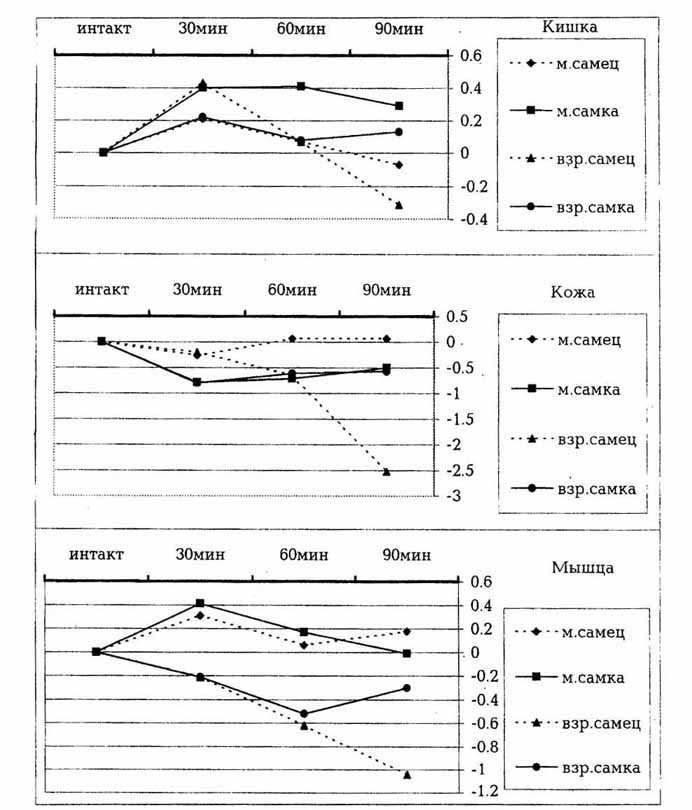
Из полученных данных по группе половозрелых
крыс видно, что гипотермический эффект, индуцированный инъекцией яда гюрзы,
в этой группе животных сильнее проявляется у самцов, нежели у самок. Так,
через 90 мин после введения яда у взрослых самцов температура скелетных
мышц понизилась на 1,04, а у взрослых самок - на 0,3°C.
Температура артериальных сосудов понизилась на 2,52, а у самок - на 0,57°C.
Температура ободочной кишки взрослых самцов через 30 мин повысилась на
0,43°C, а затем понизилась и к 90
мин упала на 0,31°C. У взрослых самок
изменения аналогичных параметров происходили менее выраженно: температура
повышалась на 0,22 и далее снижалась на 0,13°C.
Изменения температуры исследуемых областей у опытных молодых животных менее
выражены по сравнению с группой половозрелых особей. Так, у молодых самцов
через 30 мин после действия яда температура ободочной кишки повысилась
на 0.21, а у самок на 0,4°C, затем
через 90 мин понизилась на 0,07°C
у самцов и 0,29°C у самок.
|
|
Изменение температуры различных областей тела у молодых
(м) и
Температура скелетных мышц у самок после первоначального
повышения понизилась быстрее, и через 90 мин разница составила -0,01°C,
в то время как у самцов изменение составило +0,18°C.
Институт физиологии им.Л.А. Орбели НАН РА
Литература 1. Артемов Н.М., Орлов Б.Н. В сб.: Мат. XXI
Междунар. конф. по пчеловодству. М., 1967.
Температура кожи над хвостовой артерией
через 30 мин после введения яда у молодых самок упала на -0,79,
а у самцов - на -0,26°C.
Через 90 мин изменения составили соответственно -0,5
и +0,07°C. На основании опытных данных
построены графики изменения температуры исследуемых областей, которые показаны
на рисунке.
Экспериментальные данные однозначно указывают
на то, что наиболее чувствительными к яду гюрзы оказались самцы половозрелых
крыс. Температура как ободочной кишки, так и бедренной мышцы и кожи у зрелых
самцов претерпела наиболее выраженную гипотермию по сравнению с тремя другими
группами - половозрелыми самками, молодыми самцами и молодыми самками.
Введение яда гюрзы обычно приводит к начальному свертыванию крови и тромбированию,
которое происходит во всем организме, т. е. является диссеминированным,
что приводит к значительной ишемизации тканей различных органов. Тромбирование
провоцирует образование пептидаз для их рассасывания, вследствие чего возникает
спазм сосудов, что еще больше усиливает ишемизацию различных органов. К
спазму сосудов приводит также активизация симпато-адреналовой системы [9].
Можно предположить, что терморегуляционный
ресурс, базируемый на лабильности симпатической нервной системы, обеспечивающий
температурный гомеостаз организма, истощается быстрее всего у взрослых
самцов. Видимо, зрелые самцы, с одной стороны, не обладают той лабильностью
симпатической системы, которая присуща молодым особям, а с другой стороны,
не имеют того нервно-гемодинамического компенсационного ресурса, который
наличествует у взрослых самок. Этот факт говорит о том, что взрослые самцы
крыс могут быть отнесены к наиболее маргинальной группе из всех исследованных
групп по развитию терморегуляторных механизмов организма.
2. Орлов Б.Н., Вальцева И.А. Яды змей. Ташкент:
Медицина, 1977. 251 с.
3. Кононенко И.Ф. В сб.: Мат. XIX Междунар.
конф. по пчеловодству. М., 1958.
4. Арутюнян Р.А., Арутюнян К.Р., Мартиросян
С.Ш., Восканян А.В.- ДНАН Армении. 1998. Т.98. № 2. С. 175-178.
5. Султанов М.Н. Лечебные свойства змеиного
и пчелиного ядов. Ашхабад: Туркменистан, 1972. 125 с.
6. Хомутов А.Е., Дерюгина А.В., Зимина Т.А.
В сб.: Мат. конф. "Кислотно-основной и температурный гомеостаз: физиология,
биохимия и клиника". Сыктывкар. 1997. С. 175-178.
7. Schrocter R., Vogt W. - Arch. Pharm.
1973. № 2. P. 201-207.
8. Восканян А.В. В сб.: Науч. сессия, посвященная
50-летию организации Ин-та физиологии НАН РА. Ереван 1993.
9. Авакян О.М. Симпато-адреналовая система.
Ереван: Изд-во АН АрмССР, 1973. 256 с.