|
|
А. Р. Джербашьян, Р. А. Захарян, П. А. Казарян,
С.Н. Симонян, академик К.Г. Карагезян
Действие дсРНК и полинуклеотидов на апоптоз
(Представлено 19/VII 1999)
Индукция глюкокортикоидами межнуклеосомной
фрагментации ДНК хроматина впервые была показана в [1]. В дальнейшем описанное
явление получило название "запрограммированной гибели" клетки. Это феномен,
являющийся наиболее характерным биохимическим тестом апоптоза [2]. Апоптоз
рассматривается как необходимый процесс для поддержания гоместаза организма
(3). Известны гены как индуцирующие (ced 3, ced 4), так и подавляющие (red
9) апоптоз [4].
Показано, что двухспиральная РНК (дсРНК)
обладает антивирусной, интерферониндуци-рующей, противоопухолевой и иммуностимулирующей
активностью [5, 6].
В данной работе мы исследовали действие
дсРНК, полинуклеотидов поли (А), поли (У) и поли (Г) на апоптоз как с целью
изучения регуляции апоптоза, так и для оценки антивирусного и противоопухолевого
действия вышеуказанных веществ.
В работе использовали 3 - 4-недельные белые
крысы. Клетки тимуса извлекали гомогенизацией тимуса в слабо притертом
гомогенизаторе Даунса в среде RPMI 1640, 2 мМ глютамин с 10% телячьей эмбриональной
сывороткой. Суспензию клеток фильтровали через 4 слоя марли и инкубировали
4 ч при 37°С.
После инкубации клетки осаждали центрифугированием
400 х g 10 мин. Осадок клеток суспензировали в 20 мМ трис-HCl рН 7.4, 25
мм Na-ЭДТА буфере, клетки лизировали добавлением тритона Х-100 до конечной
концентрации 1% с последующей инкубацией 10 мин при 0°С. Добавляли Na-додецилсульфат
до конечной концентрации 0.5%. ДНК выделяли обработкой протеиназой К, депротенизацией
смесью хлороформ - изоамиловым спиртом (24:1) и последующей обработкой
РНК-азой.
Фрагменты ДНК разделяли электрофоретически
в 1.5%-ной агарозе в системе трис-борат рН 8.0 [7].
In vitro инкубация тимоцитов без добавок
вызывает слабый апоптоз. В наших экспериментах мы обнаружили слабую межнуклеосомную
деградацию ДНК хроматина при инкубации тимоцитов в среде без добавок (рис.
1, ряд 1).
|
|
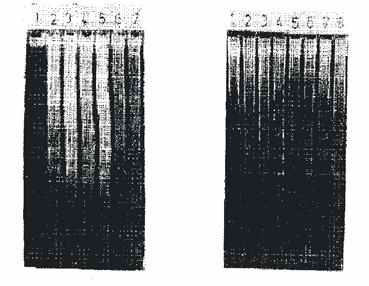
Рис.1. Рис.2.
Рис.1. Действие дсРНК на межнуклесомную фрагментацию
ДНК хромaтина тимоцитов крыс: 1-контроль без добавок; 2-инкубация клеток
с дексаметазон ацетатом 5 мкг/мл; 3-инкубация с дексаметазон ацетатом 10
мкг/мл; 4-инкубация с дексаметазон ацетатом 10-6
м; 5-инкубация с дсРНК 100 мкг/мл; 6-инкубация с дсРНК 250 мкг/мл; 7-инкубация
с дсРНК 500 мкг/мл; 8-инкубация с дсРНК 250 мкг/мл и дексаметазон ацетатом
10-6 м.
Рис.2. Действие полинуклеотидов на межнуклесомную
фрагментацию ДНК хромaтина тимоцитов: 1-ДНК тимуса без инкубации, контроль;
2-инкубация клеток с поли (А) 50 мкг/мл; 3-инкубация с поли (А) 100 мкг/мл;
4-инкубация с поли (У) 50 мкг/мл; 5-инкубация с поли (У) 100 мкг/мл; 6-инкубация
с
поли (Г) 50 мкг/мл; 3-инкубация с поли (Г) 100 мкг/мл.
дсРНК индуцирует апоптоз при in vitro действии
и в концентрациях 100, 250 и 500 мкг/мл вызывает межнуклеосомную фрагментацию
ДНК хроматина тимоцитов (рис. 1, ряд 5-7).
В качестве контрольного апоптоза анализировали
действие дексаметазонацетата (рис. 1, ряд 2-4). Индукция апоптоза усиливается
при совместном действии дсРНК и дексаметазон ацетата (рис. 1, ряд 8).
Полинуклеотиды поли (А), поли (У) и поли
(Г) индуцируют апоптоз и в концентрациях 50 и 100 мкг/мл вызывают межнуклесомальную
фрагментацию хроматина (рис. 2, ряды 2-7; ряд 1 - ДНК тимоцитов без инкубации).
Наибольший индуцирующий эффект на апоптоз
имеет действие поли (У), наименьший индуцирующий эффект имеет поли (Г).
Ранее было показано лигандообразование дсРНК
с белками мозга [8].
Известно, что при вирусных инфекциях и раковых
образованиях подавляется апоптоз [3, 9].
Полученные результаты показали, что молекулярный
механизм действия дсРНК, поли (А), поли (У) и поли (Г) опосредован через
индукцию запрограммированной гибели клетки.
Вышеуказанные вещества, а из полинуклеотидов
особенно поли (У), могут быть рекомендованы к применению при вирусных заболеваниях
как апоптоз - индуцирующие вспомогательные терапевтические средства.
Институт молекулярной
биологии НАН РА
Республиканский гематологический
центр
Литература
1. Захарян Р.А., Погосян Р.Г.ДАН АрмССР 1978.
T. 67. № 2. С. 110-113.
2. Willie A.N. - Nature. 1980. V. 284. P.
555-556.
3. Thompson C.B. - Science. 1995. V. 267.
P. 1456-1462.
4. Steller H. - Sciense - 1995.V. 267. P.
1445-1449.
5. Фелдмане Г.Я., Умбрашко Ю.Б., Буйкис
А.Х. и др. - Вопр. вирусологии.1984. Т. 4. С. 463-468.
6. Фомина А.Н., Григорян С.С., Зайцева О.В.
и др. - Антибиотики.1980.Т. 1. С. 28-32.
7. Хансон К.П. - Изв. АН РФ. Сер. биол.1998.
Т. 2. С. 134-141.
8. Маниатис Т., Фриг Э., Сембрук Дж. Методы
генетической инженерии. М.,1984.
9. Zakharyan R.A. - QLT/congress, ENA/FBBS-BRA.
Cambridge. 1991.